

Regeneration of broken tip links and restoration of mechanical transduction in hair cells
- PMID: 8986835
- PMCID: PMC26428
- DOI: 10.1073/pnas.93.26.15469
Regeneration of broken tip links and restoration of mechanical transduction in hair cells
Abstract
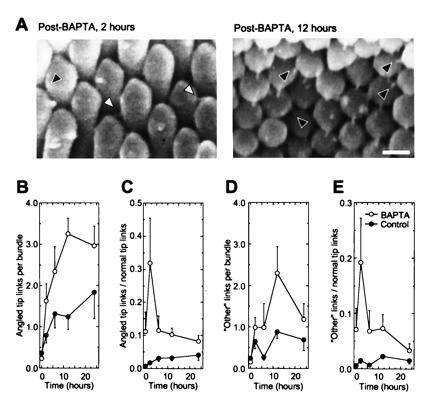
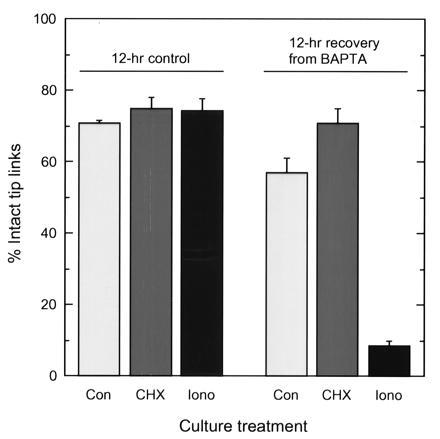
A hair cell's tip links are thought to gate mechanoelectrical transduction channels. The susceptibility of tip links to acoustic trauma raises questions as to whether these fragile structures can be regenerated. We broke tip links with the calcium chelator 1,2-bis(O-aminophenoxy) ethane-N,N,N',N'-tetraacetic acid and found that they can regenerate, albeit imperfectly, over several hours. The time course of tip-link regeneration suggests that this process may underlie recovery from temporary threshold shifts induced by noise exposure. Cycloheximide does not block tip-link regeneration, indicating that new protein synthesis is not required. The calcium ionophore ionomycin prevents regeneration, suggesting regeneration normally may be stimulated by the reduction in stereociliary Ca2+ when gating springs rupture and transduction channels close. Supporting the equivalence of tip links with gating springs, mechanoelectrical transduction returns over the same time period as tip links; strikingly, adaptation is substantially reduced, even 24 hr after breaking tip links.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
