

A direct comparison of the single-channel properties of synaptic and extrasynaptic NMDA receptors
- PMID: 8987740
- PMCID: PMC6793703
- DOI: 10.1523/JNEUROSCI.17-01-00107.1997
A direct comparison of the single-channel properties of synaptic and extrasynaptic NMDA receptors
Abstract
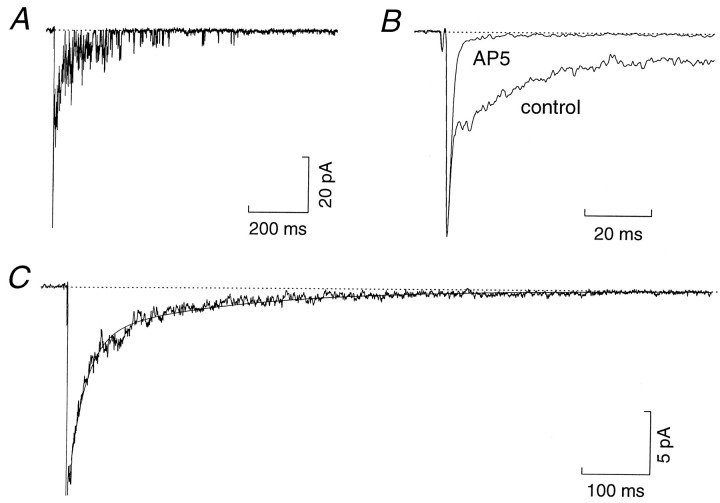
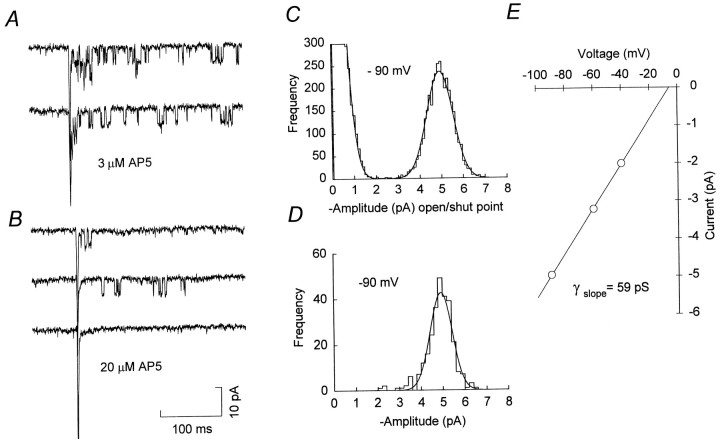
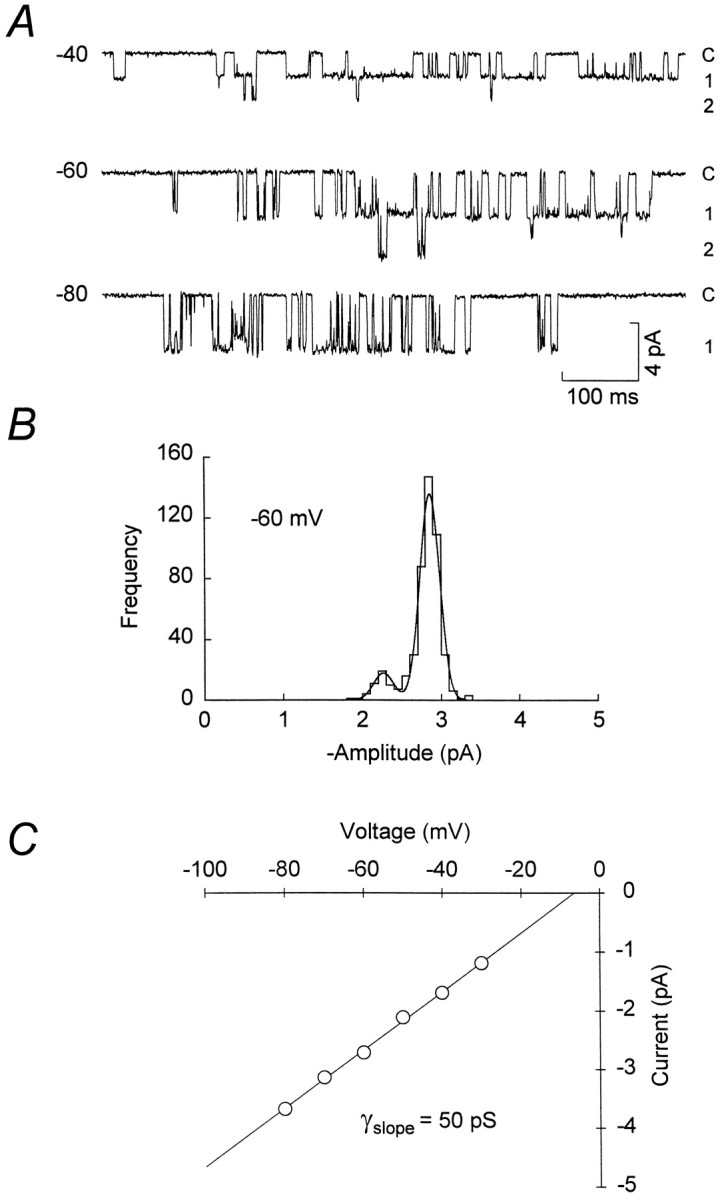
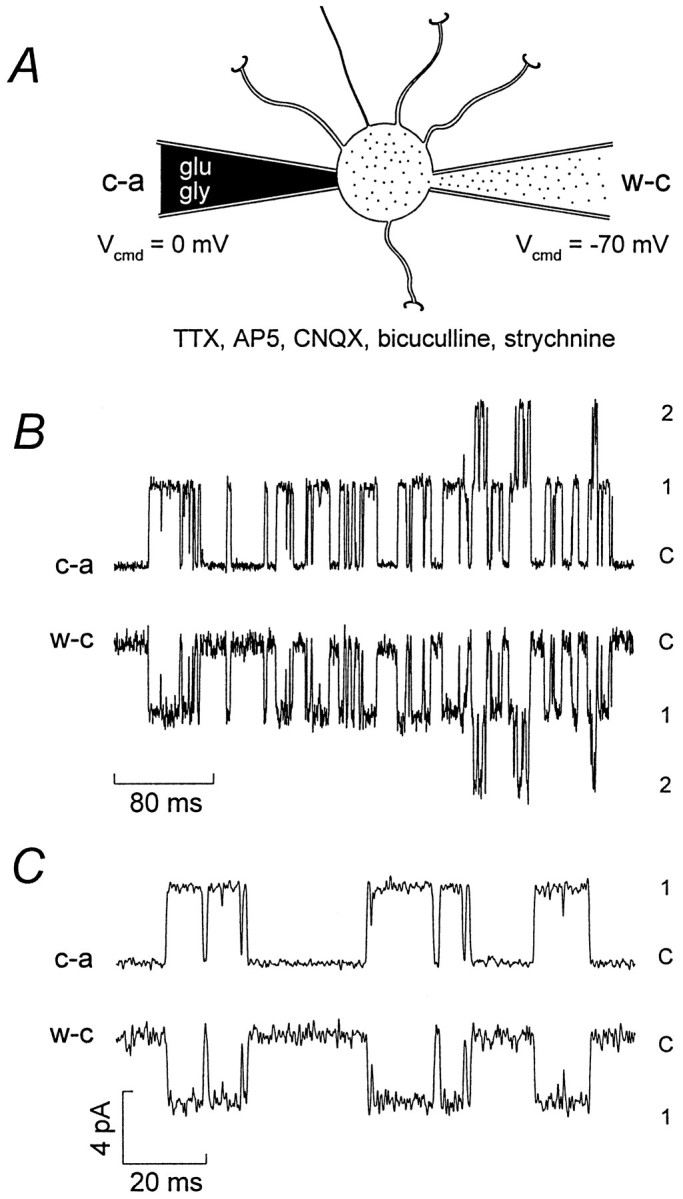
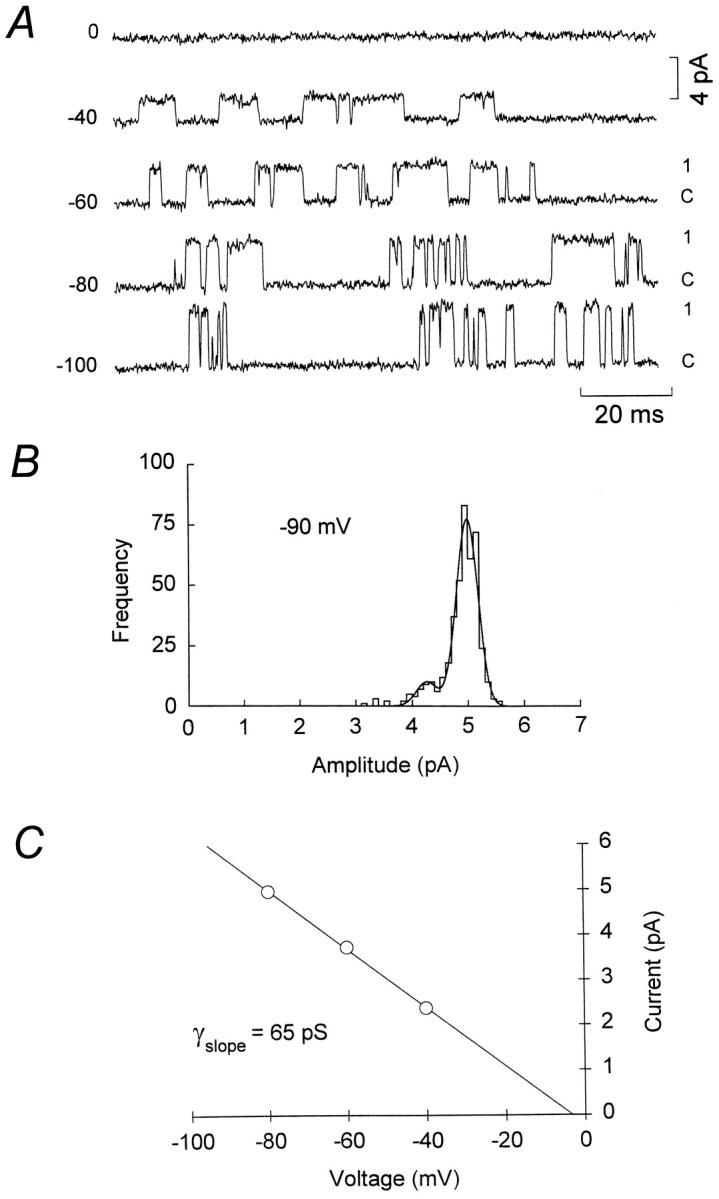
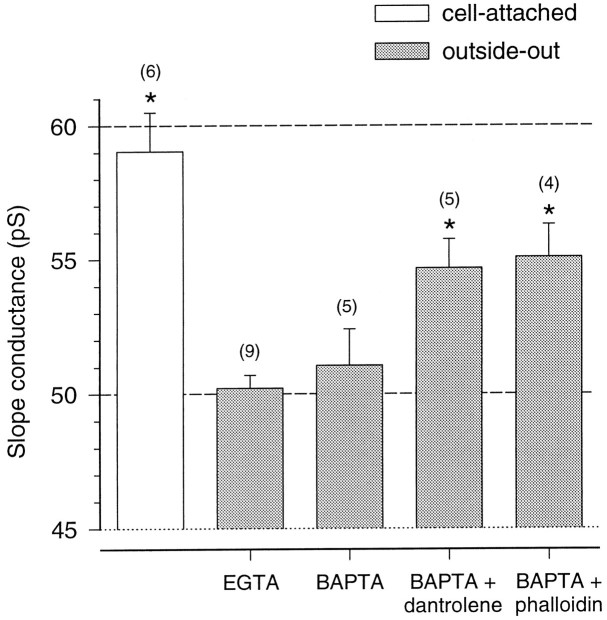
The assumption that synaptic and extrasynaptic glutamate receptors are similar underpins many studies that have sought to relate the behavior of channels in excised patches to the macroscopic properties of the EPSC. We have examined this issue for NMDA receptors in cerebellar granule cells, the small size of which allows the opening of individual synaptic NMDA channels to be resolved directly. We have used whole-cell patch-clamp recordings to determine the conductance and open time of NMDA channels activated during the EPSC and used cell-attached and outside-out recordings to examine NMDA receptors in somatic membrane. Conductance and open time of synaptic channels were indistinguishable from those of extrasynaptic channels in cell-attached patches. However, the channel conductance in outside-out patches was 20% lower than in cell-attached recordings. This change was partially reduced by dantrolene and phalloidin, suggesting that it may involve depolymerization of actin following Ca2+ release from intracellular stores. Our results demonstrate that synaptic and extrasynaptic NMDA receptors have similar microscopic properties. However, NMDA channel conductance is reduced following the formation of an outside-out patch.
Figures
References
-
- Akazawa C, Shigemoto R, Bessho Y, Nakanishi S, Mizuno N. Differential expression of five N-methyl-d-aspartate receptor subunit mRNAs in the cerebellum of developing and adult rats. J Comp Neurol. 1994;347:150–160. - PubMed
-
- Barry PH, Lynch JW. Liquid junction potentials and small cell effects in patch-clamp analysis. J Membr Biol. 1991;121:101–117. - PubMed
-
- Bekkers JM, Stevens CF. NMDA receptors at excitatory synapses in the hippocampus: test of a theory of magnesium block. Neurosci Lett. 1993;156:73–77. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous