

Synaptic integration of functionally diverse pallidal information in the entopeduncular nucleus and subthalamic nucleus in the rat
- PMID: 8987757
- PMCID: PMC6793683
- DOI: 10.1523/JNEUROSCI.17-01-00308.1997
Synaptic integration of functionally diverse pallidal information in the entopeduncular nucleus and subthalamic nucleus in the rat
Abstract
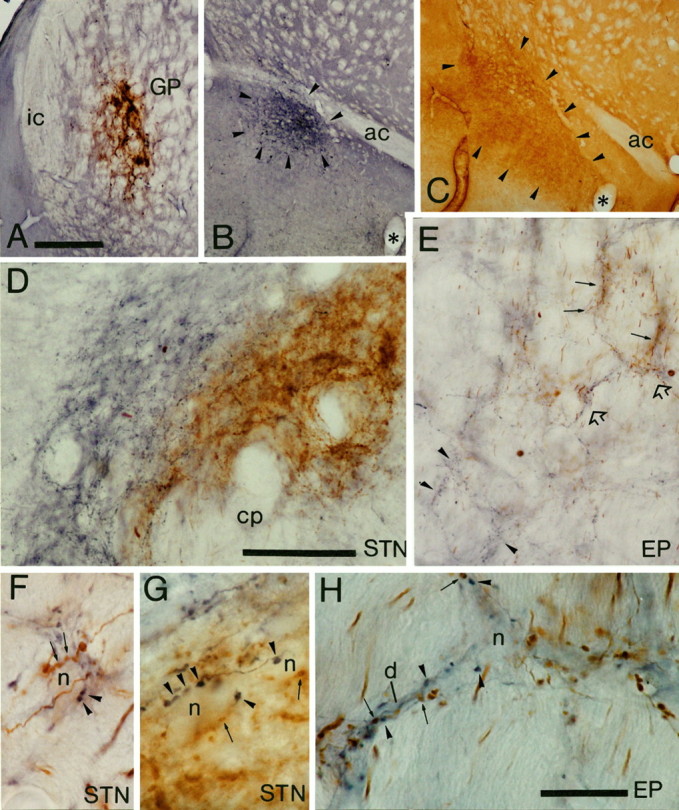
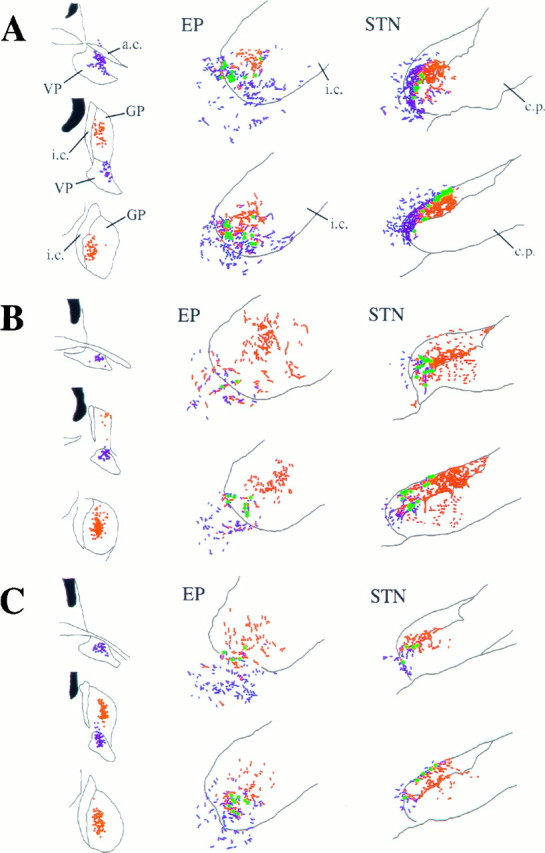
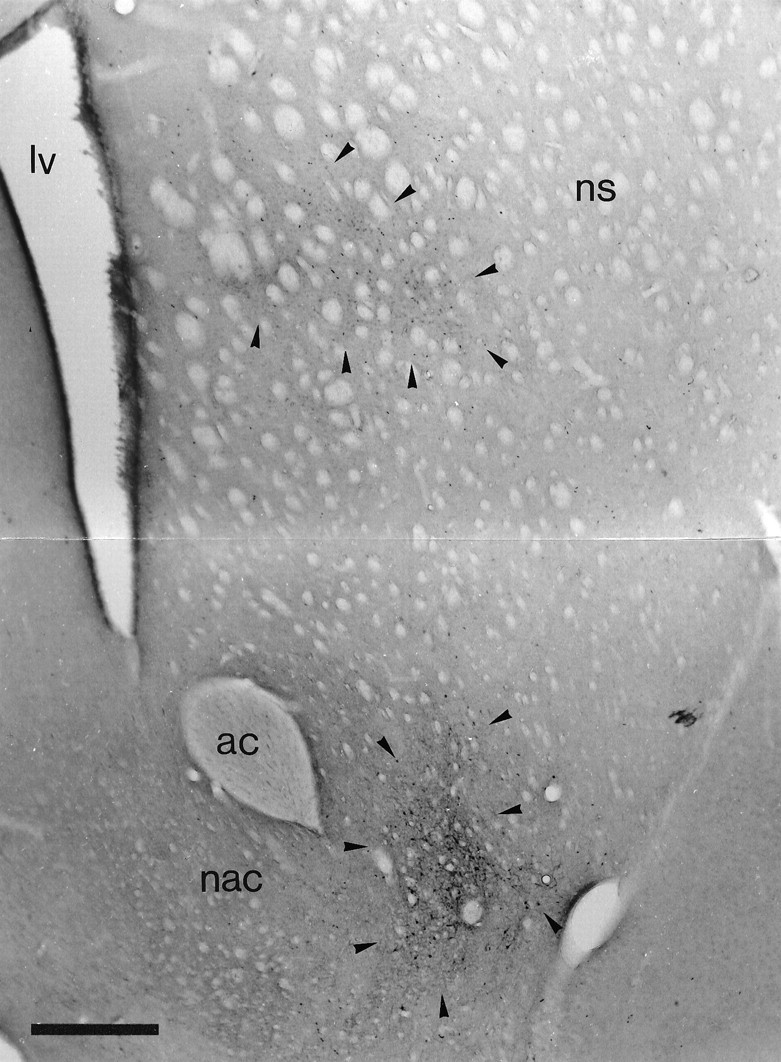
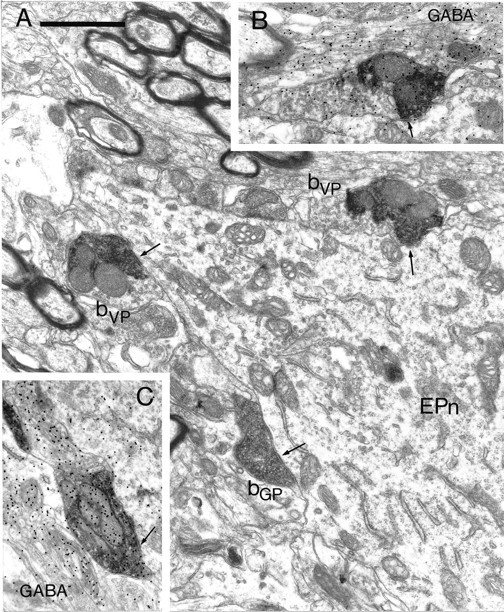
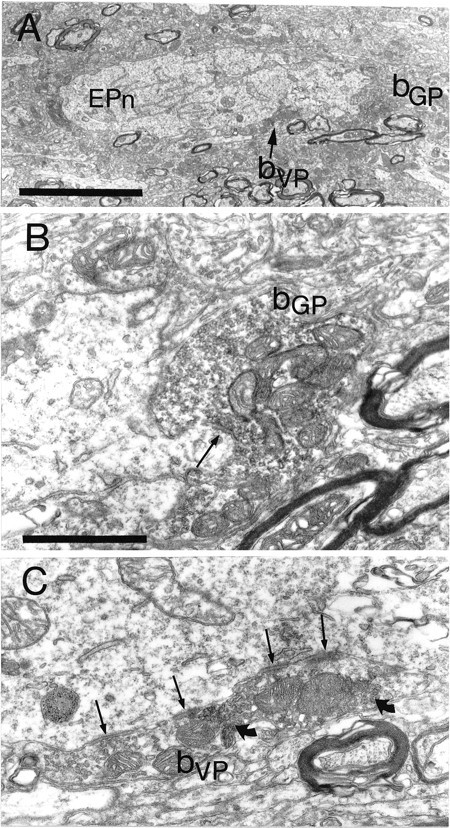
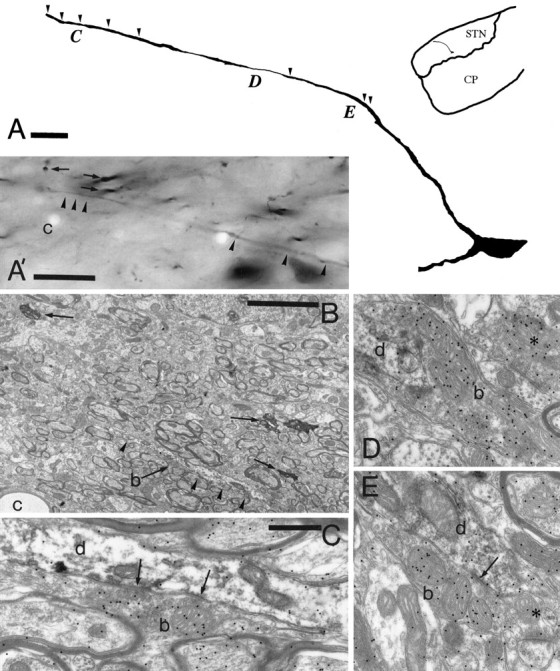
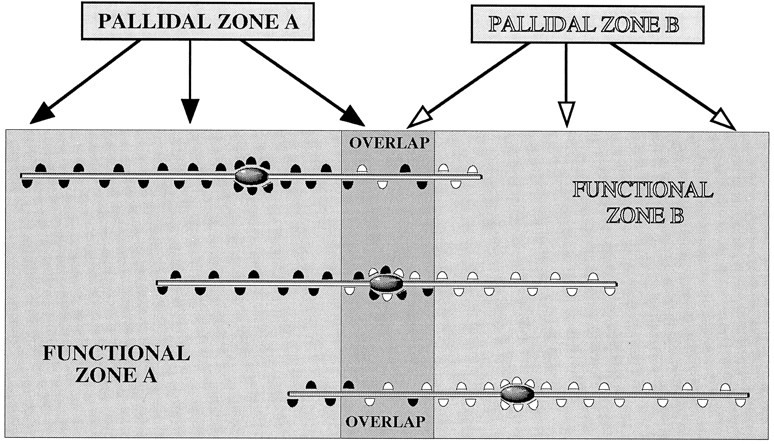
To determine the principles of synaptic innervation of neurons in the entopeduncular nucleus and subthalamic nucleus by neurons of functionally distinct regions of the pallidal complex, double anterograde labeling was carried out at both light and electron microscopic levels in the rat. Deposits of the anterograde tracers Phaseolus vulgaris-leucoagglutinin and biotinylated dextran amine were placed in different functional domains of the pallidal complex in the same animals. The tracer deposits in the ventral pallidum and the globus pallidus gave rise to GABA-immunopositive projections to the entopeduncular nucleus, the subthalamic nucleus, and the more medial lateral hypothalamus that were largely segregated but overlapped at the interface between the two fields of projection. In these regions the proximal parts of individual neurons in the entopeduncular nucleus, lateral hypothalamus, and subthalamic nucleus received synaptic input from terminals derived from both the ventral pallidum and the globus pallidus. Furthermore, the analysis of the afferent synaptic input to the dendrites of neurons in the subthalamic nucleus that cross functional boundaries of the nucleus defined by the pallidal inputs, revealed that terminals with the morphological and neurochemical characteristics of those derived from the pallidal complex make synaptic contact with all parts of the dendritic tree, including distal regions. It is concluded that functionally diverse information carried by the descending projections of the pallidal complex is synaptically integrated by neurons of the entopeduncular nucleus, lateral hypothalamus, and subthalamic nucleus by two mechanisms. First, neurons located at the interface between functionally distinct, but topographically adjacent, projections could integrate diverse information by means of the synaptic convergence at the level of the cell body and proximal dendrites. Second, because the distal dendrites of neurons in the subthalamic nucleus receive input from the pallidum, those that extend across two distinct domains of pallidal input could also provide the morphological basis of integration.
Figures



References
-
- Albin RL, Young AB, Penney JB. The functional anatomy of basal ganglia disorders. Trends Neurosci. 1989;12:366–375. - PubMed
-
- Alexander GE, Crutcher MD. Functional architecture of basal ganglia circuits: neural substrates of parallel processing. Trends Neurosci. 1990;13:266–271. - PubMed
-
- Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci. 1986;9:357–381. - PubMed
-
- Alexander GE, Crutcher MD, DeLong MR. Basal ganglia-thalamocortical circuits: parallel substrates for motor, oculomotor, “prefrontal” and “limbic” functions. In: Uylings HBM, Vaneden CG, Debruin JPC, Corner MA, Feenstra MGP, editors. Prefrontal cortex. Its structure, function and pathology. Elsevier; Amsterdam: 1990. pp. 119–146. - PubMed
-
- Aosaki T, Kimura M, Graybiel AM. Temporal and spatial characteristics of tonically active neurons of the primate’s striatum. J Neurophysiol. 1995;73:1234–1252. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources