

Functional stages in the formation of human long-term motor memory
- PMID: 8987766
- PMCID: PMC6793707
- DOI: 10.1523/JNEUROSCI.17-01-00409.1997
Functional stages in the formation of human long-term motor memory
Abstract
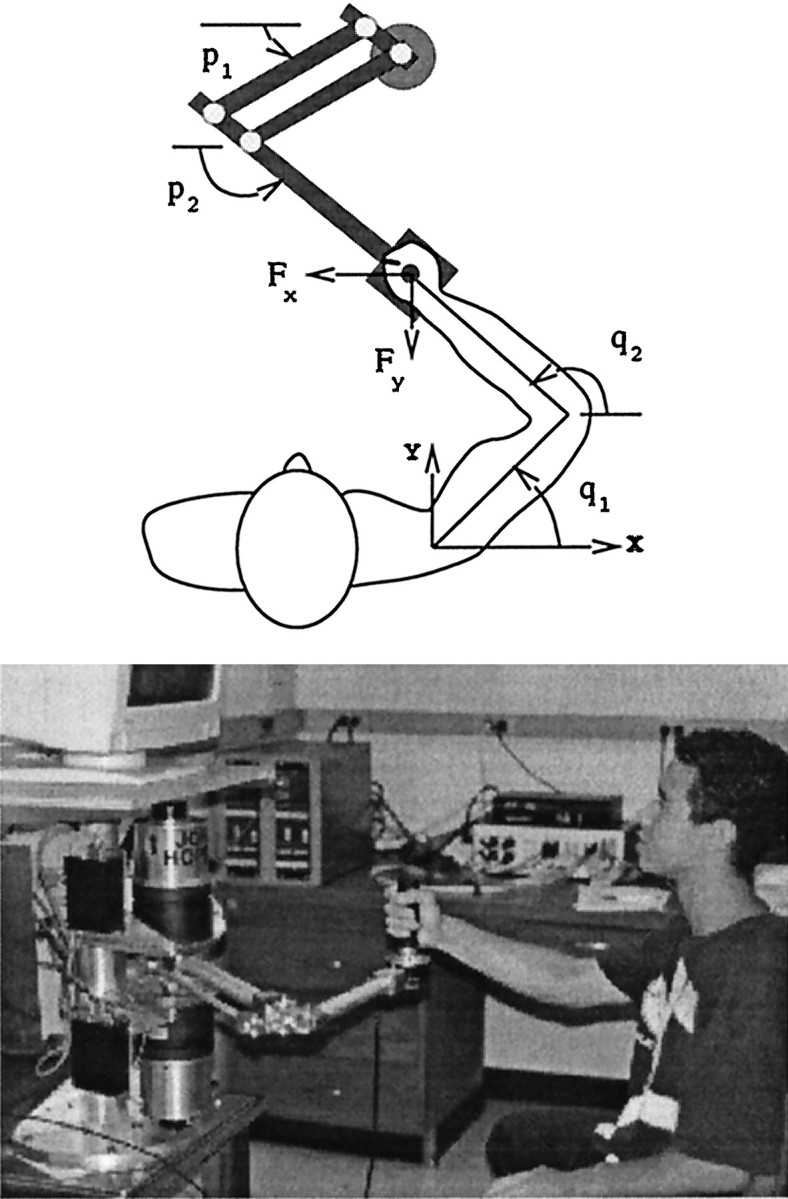
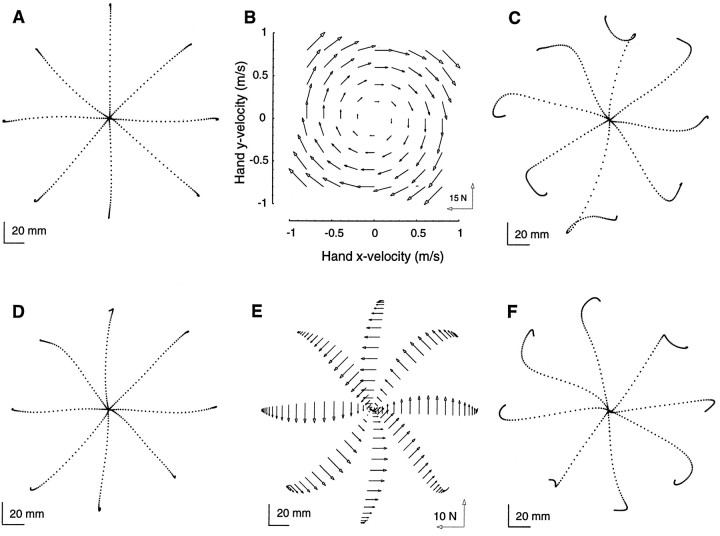
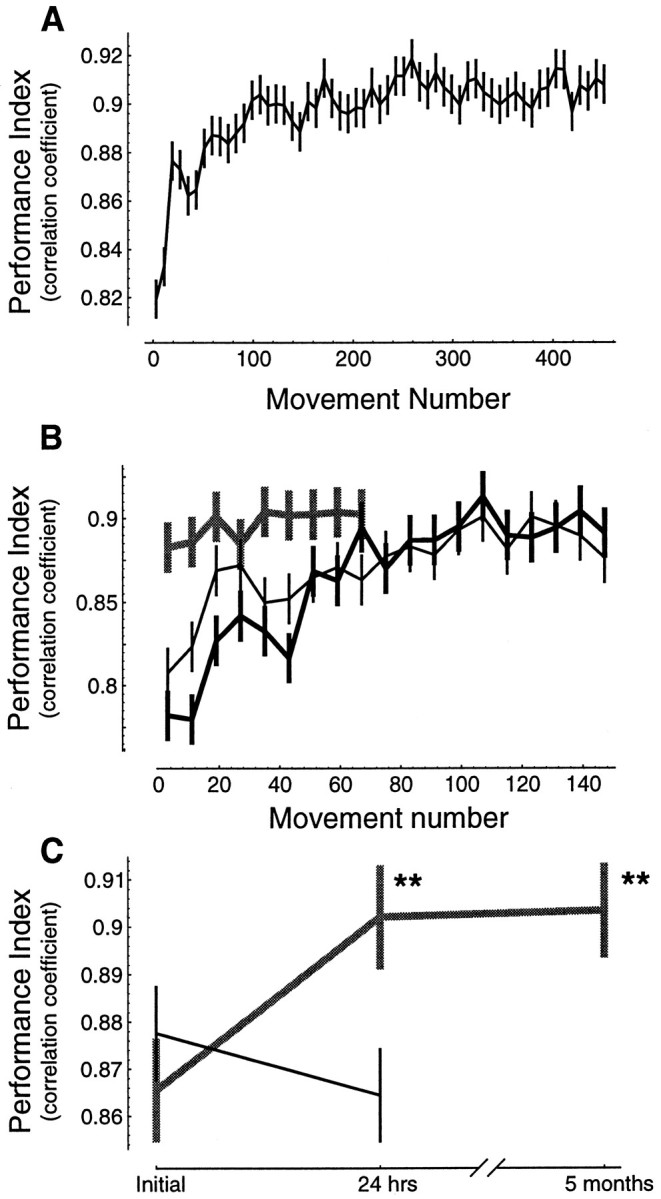
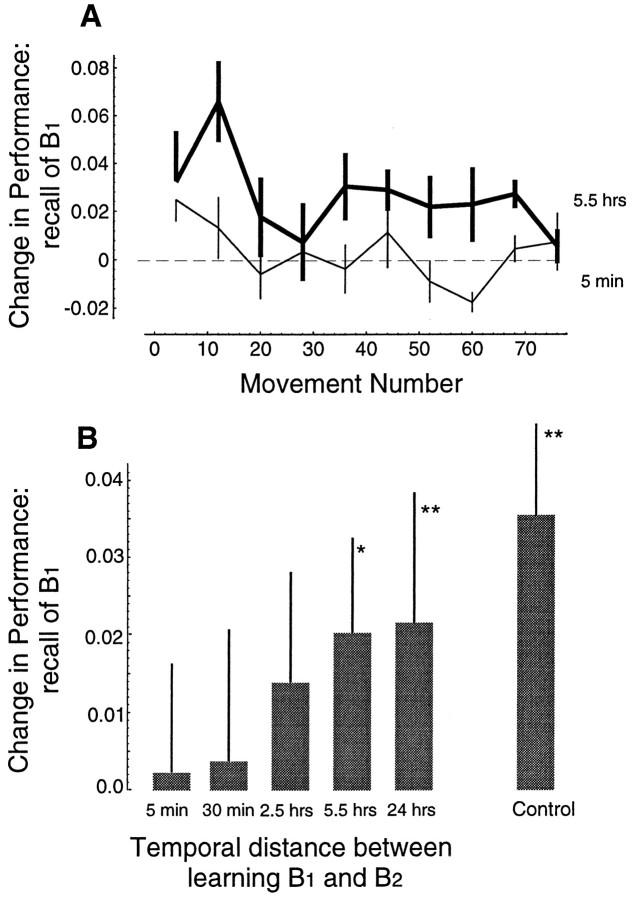
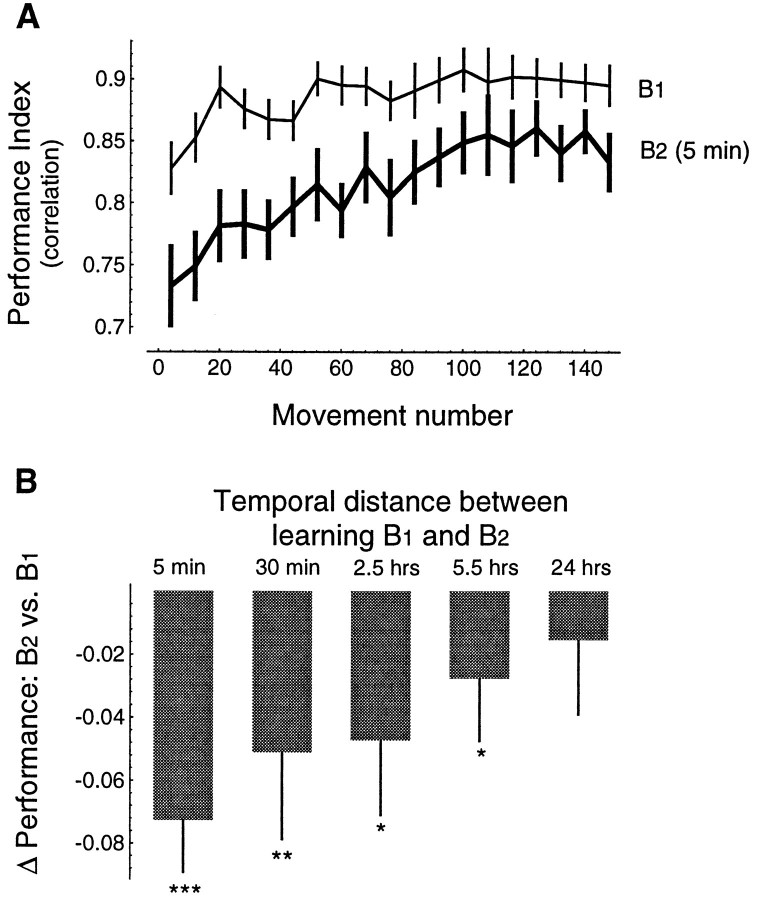
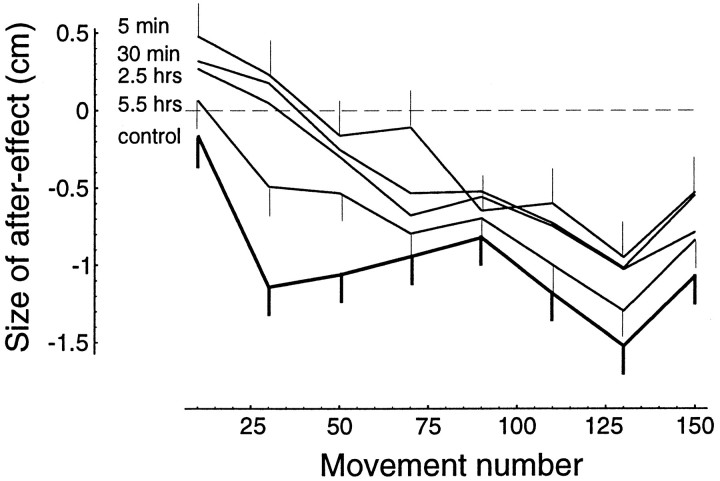
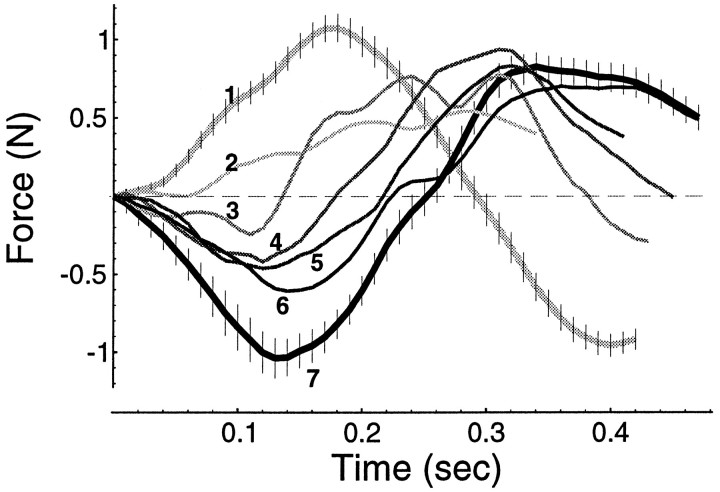
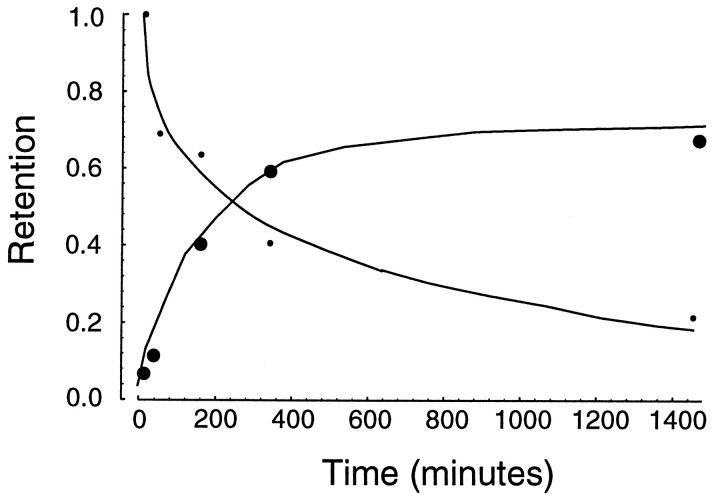
Previous research has demonstrated that the primate CNS has the ability to learn and store multiple and conflicting visuo-motor maps. Here we studied the ability of human subjects to learn to make reaching movements while interacting with one of two conflicting mechanical environments as produced by a robotic manipulandum. We demonstrate that two motor maps may be learned and retained, but only if the training sessions in the tasks are separated by an interval of approximately 5 hr. If the interval is shorter, learning of the second map begins with an internal model appropriate for the first task and performance in the second task is significantly impaired. Analysis of the after-effects suggests that with a short temporal distance, learning of the second task leads to an unlearning of the internal model for the first. With the longer temporal distance, learning of the second task starts with an unbiased internal model, and performance approaches that of naives. Furthermore, the memory of the consolidated skill lasts for at least 5 months after training. These results argue for a distinct change in the state of resistance of motor memory (to disruption) within a few hours after acquisition. We suggest that motor practice results in memories that have at least two functional components: soon after completion of practice, one component fades while another is strengthened. A further experiment suggests that the hypothetical first stage is not merely a gateway to long-term memory, but also temporary storage for items of information, whether new or old, for use in the near-term. Our results raise the possibility that there are distinct neuronal mechanisms for representation of the two functional stages of motor memory.
Figures

References
-
- Aizawa H, Inase M, Mushiake H, Shima K, Tanji J. Reorganization of activity in the supplementary motor area associated with motor learning and functional recovery. Exp Brain Res. 1991;84:668–671. - PubMed
-
- Asanuma H, Keller A. Neuronal mechanisms of motor learning in mammals. NeuroReport. 1991;2:217–224. - PubMed
-
- Bailey CH, Kandel ER. Molecular and structural mechanisms underlying long-term memory. In: Gazzaniga M, editor. The cognitive neurosciences. MIT; Cambridge, MA: 1995. pp. 19–36.
-
- Baker J, Perlmutter SO, Peterson BW, Rude SA, Robinson FR. Simultaneous opposing adaptive changes in cat vestibulo-ocular reflex direction for two body orientations. Exp Brain Res. 1987;69:220–224. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical