

Activation of hippocampal adenosine A3 receptors produces a desensitization of A1 receptor-mediated responses in rat hippocampus
- PMID: 8987783
- PMCID: PMC5470729
- DOI: 10.1523/JNEUROSCI.17-02-00607.1997
Activation of hippocampal adenosine A3 receptors produces a desensitization of A1 receptor-mediated responses in rat hippocampus
Abstract
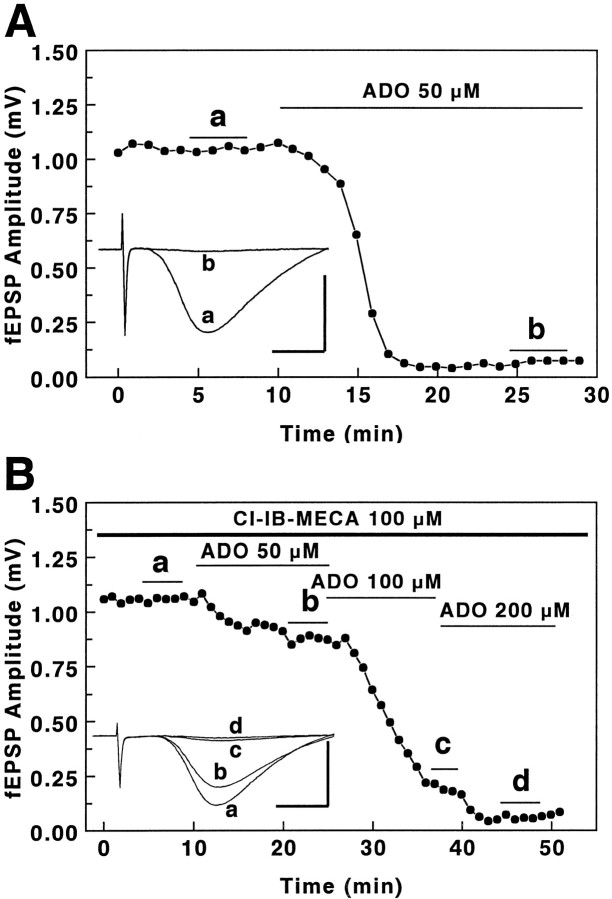
The adenosine A3 receptor is expressed in brain, but the consequences of activation of this receptor on electrophysiological activity are unknown. We have characterized the actions of a selective adenosine A3 receptor agonist, 2-chloro-N6-(3-lodobenzyl)-adenosine-5'-N-methyluronamide (Cl-IB-MECA), and a selective A3 receptor antagonist, 3-ethyl-5-benzyl-2-methyl-4-phenylethynyl-6-phenyl-1, 4-(+/-)-dihydropyridine-3,5-dicarboxylate (MRS 1191), in brain slices from rat hippocampus. In the CA1 region, activation of A3 receptors had no direct effects on synaptically evoked excitatory responses, long-term potentiation, or synaptic facilitation. However, activation of A3 receptors with Cl-IB-MECA antagonized the adenosine A1 receptor-mediated inhibition of excitatory neurotransmission. The effects of Cl-IB-MECA were blocked by pretreatment with MRS 1191, which by itself had no effect on A1 receptor-mediated responses. The presynaptic inhibitory effects of baclofen and carbachol, mediated via GABA(B) and muscarinic receptors, respectively, were unaffected by Cl-IB-MECA. The maximal response to adenosine was unchanged, suggesting that the primary effect of Cl-IB-MECA was to reduce the affinity of adenosine for the receptor rather than to uncouple it. Similar effects could be demonstrated after brief superfusion with high concentrations of adenosine itself. Under normal conditions, endogenous adenosine in brain is unlikely to affect the sensitivity of A1 receptors via this mechanism. However, when brain concentrations of adenosine are elevated (e.g., during hypoxia, ischemia, or seizures), activation of A3 receptors and subsequent heterologous desensitization of A1 receptors could occur, which might limit the cerebroprotective effects of adenosine under these conditions.
Figures








References
-
- Abbracchio MP, Brambilla R, Ceruti S, Kim HO, von Lubitz DKJE, Jacobson KA, Cattabeni F. G protein-dependent activation of phospholipase C by adenosine A3 receptors in rat brain. Mol Pharmacol. 1995;48:1038–1045. - PubMed
-
- Ali H, Cunha-Melo JR, Saul WF, Beaven MA. Activation of phospholipase C via adenosine receptors provides synergistic signals for secretion in antigen-stimulated RBL-2H3 cells. Evidence for a novel adenosine receptor. J Biol Chem. 1990;265:745–753. - PubMed
-
- Cunha RA, Constantino MD, Sebastião AM, Ribeiro JA. Modification of A1 and A2a adenosine receptor binding in aged striatum, hippocampus and cortex of the rat. NeuroReport. 1995;6:1583–1588. - PubMed
-
- Cunha RA, Johansson B, Constantino D, Sebastião AM, Fredholm BB. Evidence for high affinity binding sites for the adenosine A2A receptor agonist [3H]CGS21680 in the rat hippocampus and cerebral cortex that are different from striatal A2A receptors. Naunyn Schmiedebergs Arch Pharmacol. 1996;353:261–271. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous