

Neuronal adaptations to changes in the social dominance status of crayfish
- PMID: 8987791
- PMCID: PMC6573243
- DOI: 10.1523/JNEUROSCI.17-02-00697.1997
Neuronal adaptations to changes in the social dominance status of crayfish
Abstract
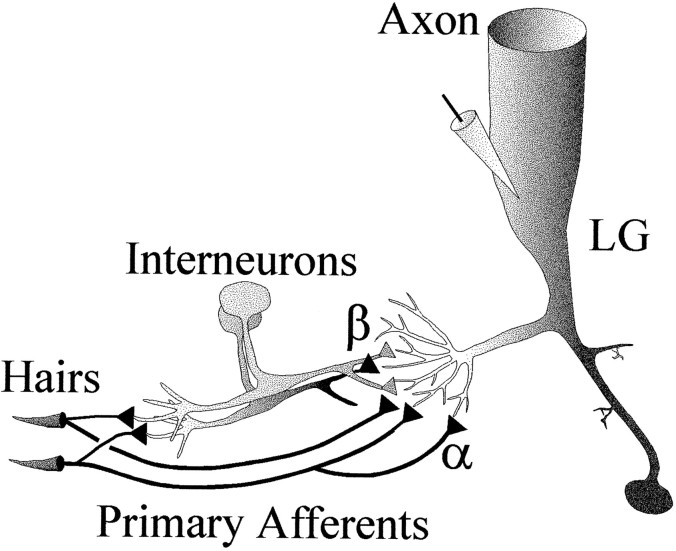
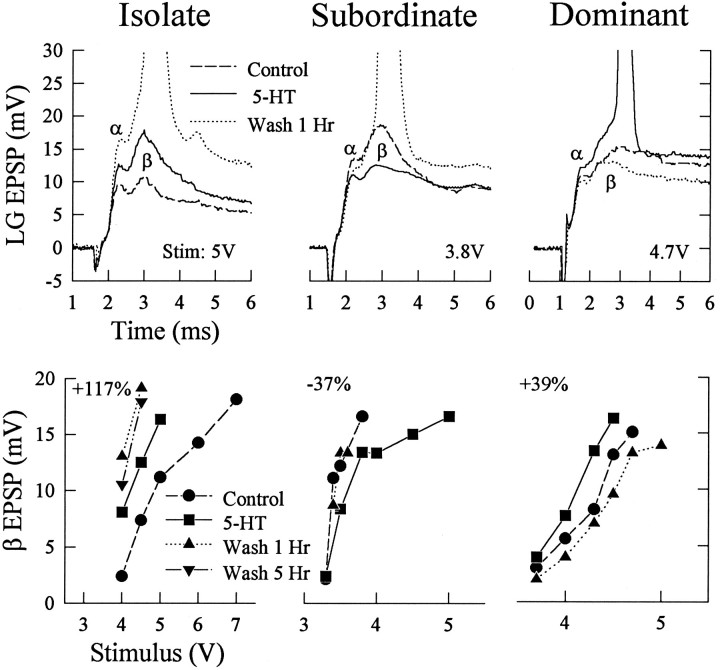
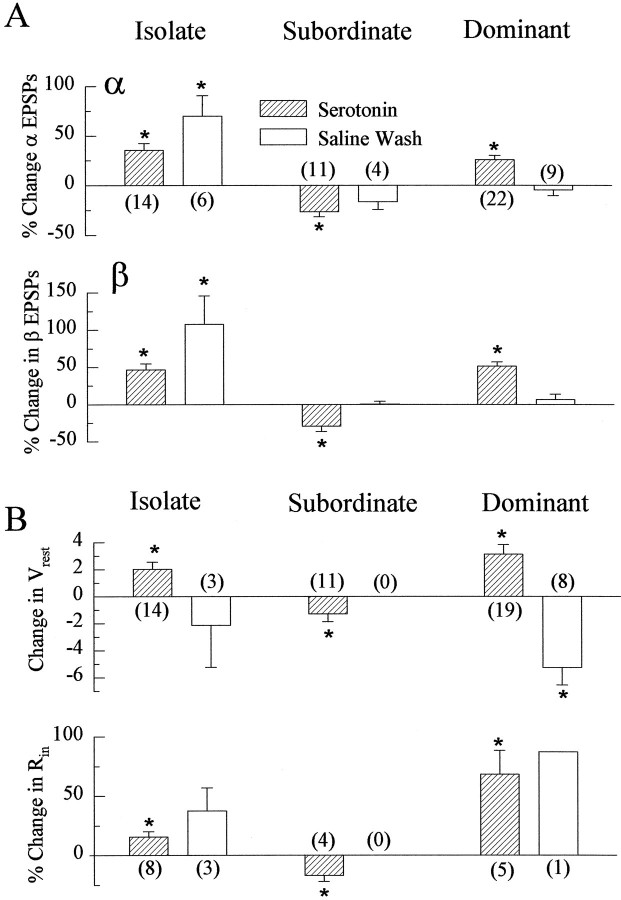
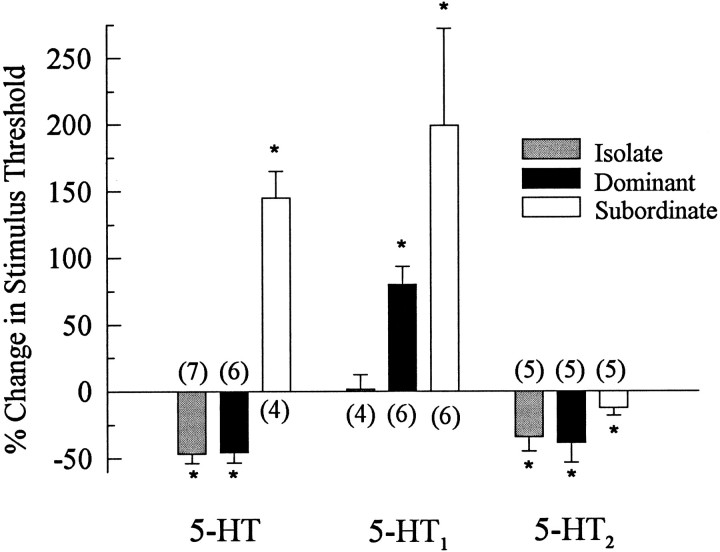
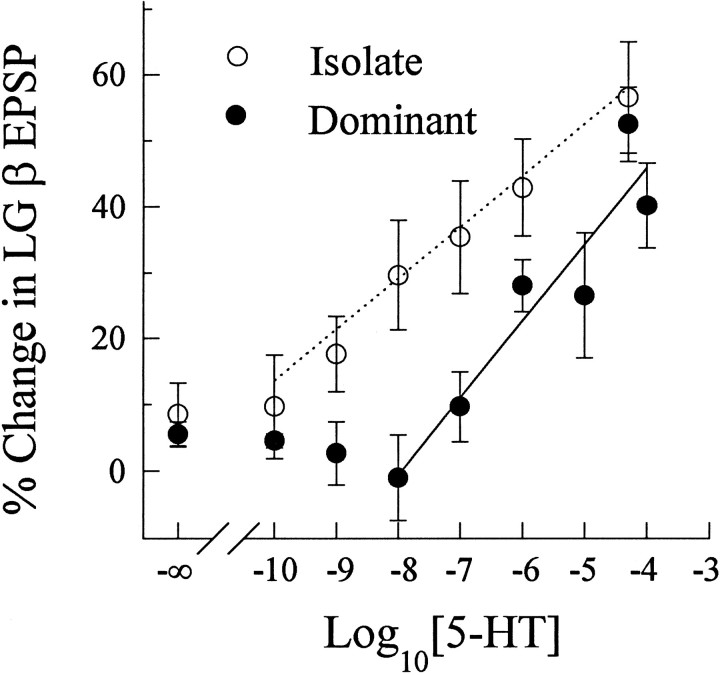
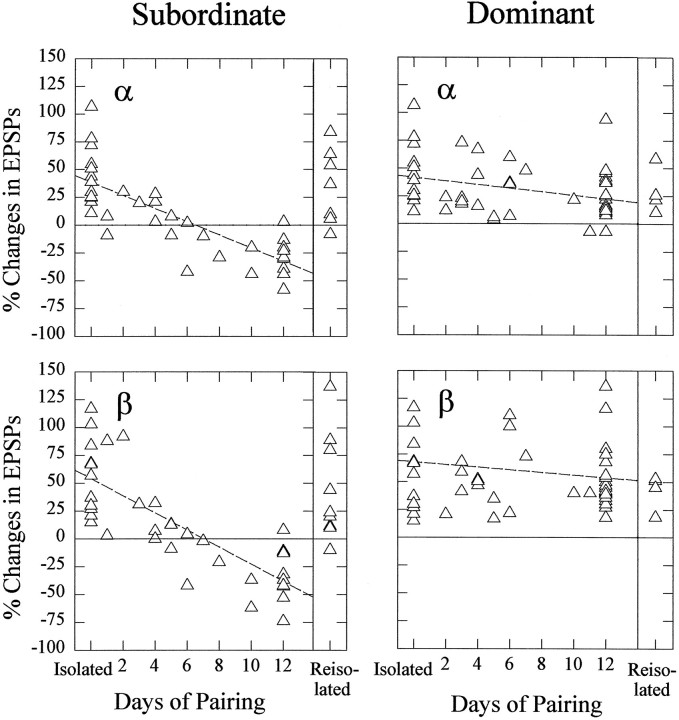
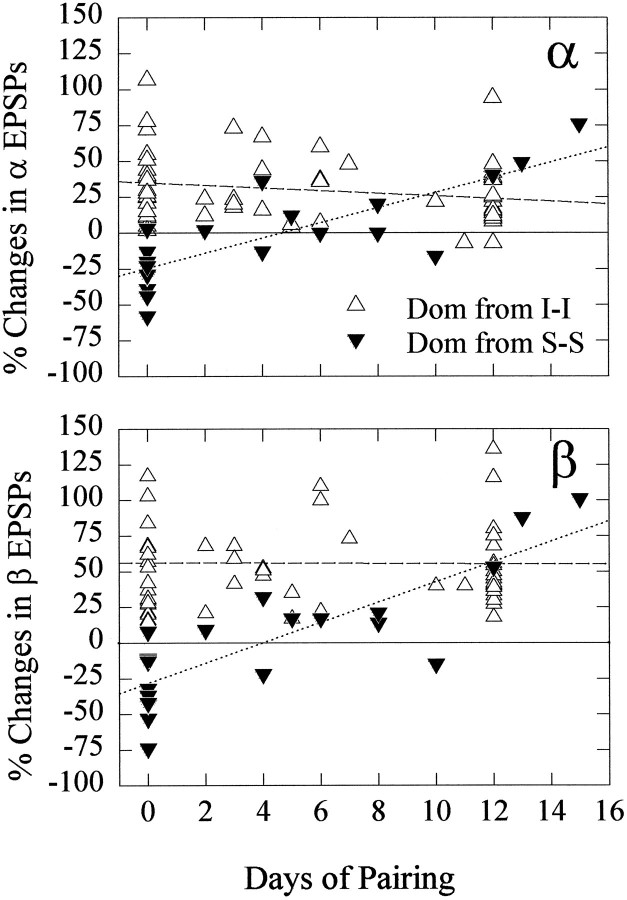
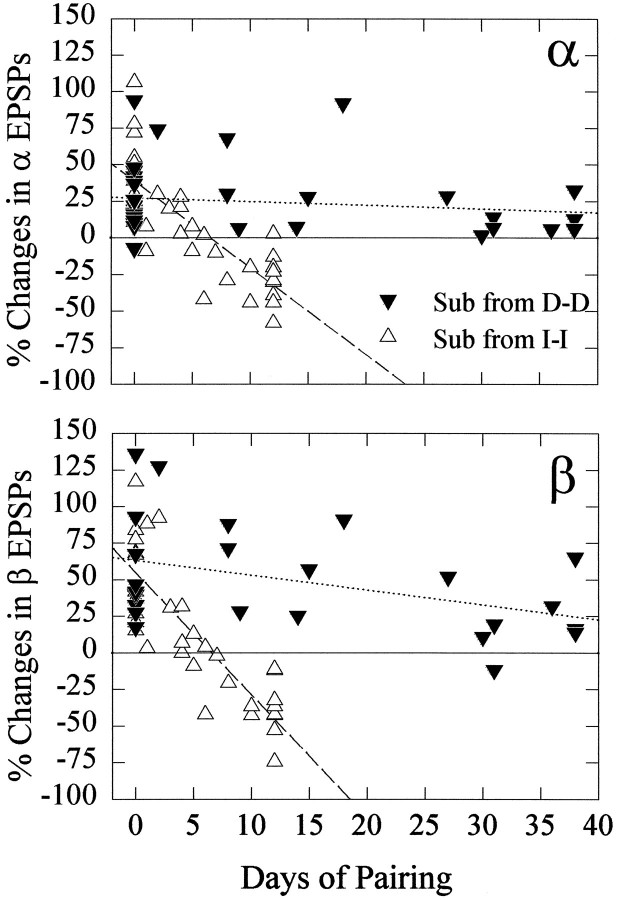
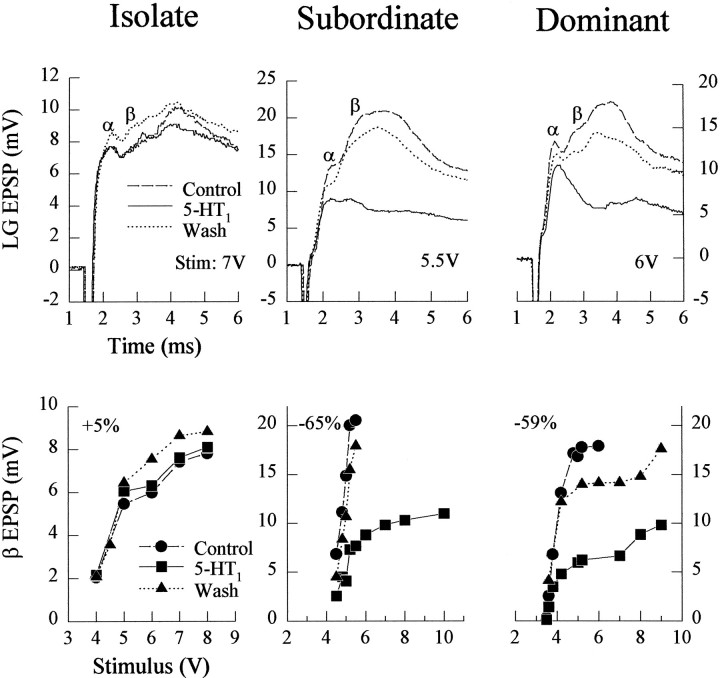
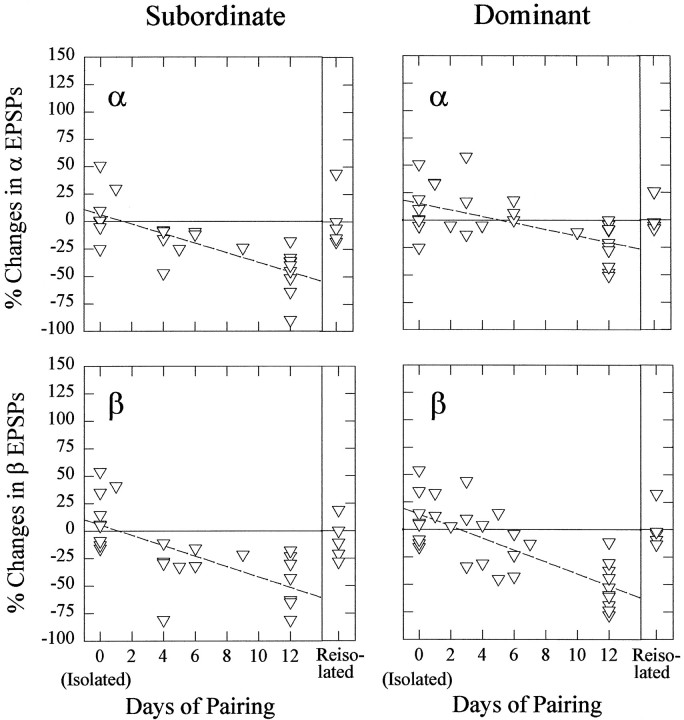
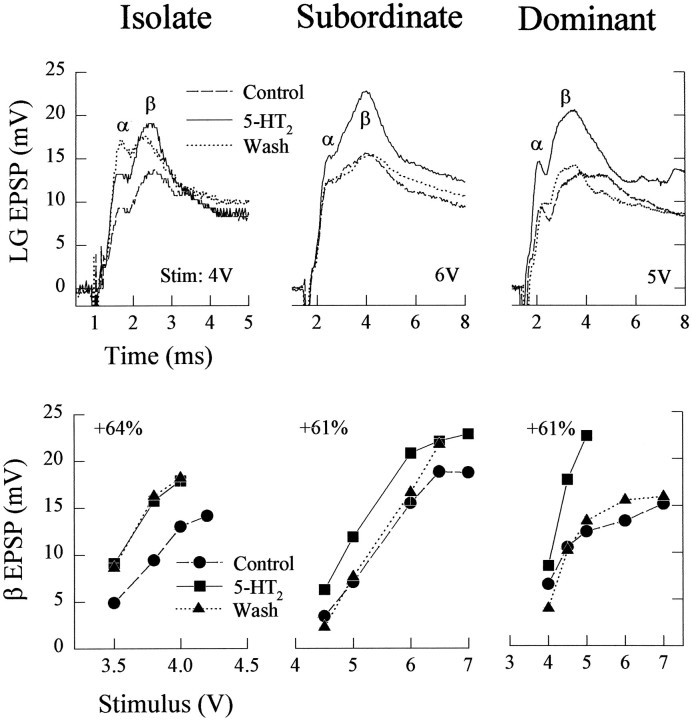
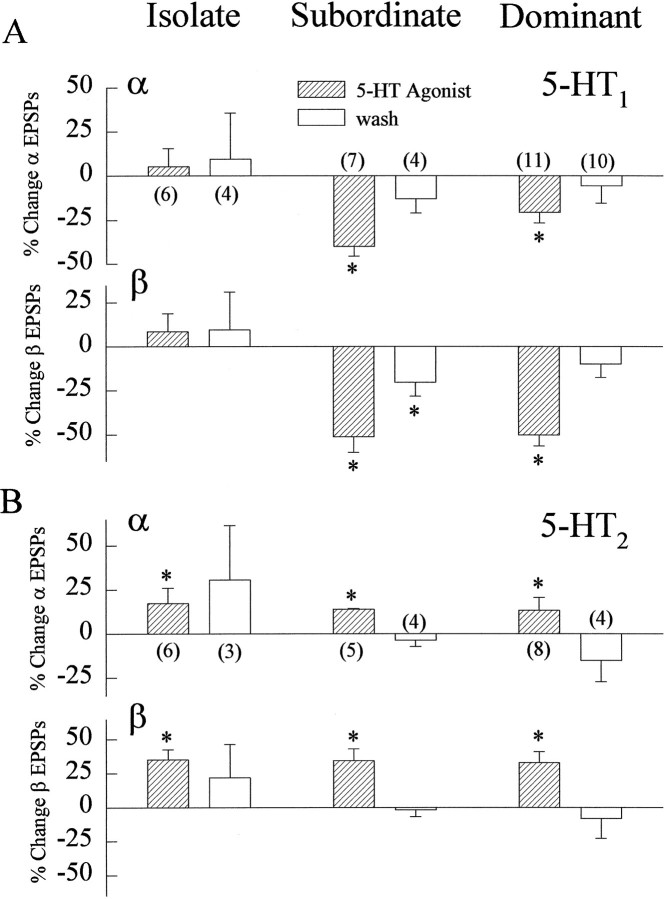
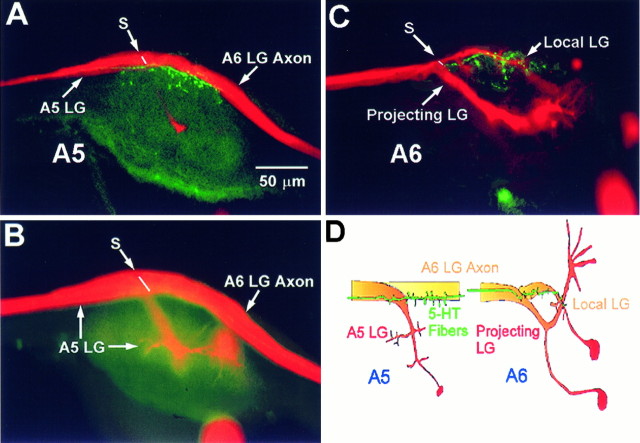
The effect of superfused serotonin (5-HT; 50 microns) on the synaptic responses of the lateral giant (LG) interneuron in crayfish was found to depend on the social status of the animal. In socially isolated animals. 5-HT persistently increased the response of LG to sensory nerve shock. After social isolates were paired in a small cage, they fought and determined their dominant and subordinate status. After 12 d of pairing, 5-HT reversibly inhibited the response of LG in the social subordinate and reversibly increased the response of LG in the social dominant crayfish. The effect of 5-HT changed approximately linearly from response enhancement to inhibition in the new subordinate over the 12 d of pairing. If, after 12 d pairing, the subordinate was reisolated for 8 d, the response enhancement was restored. If the subordinate, instead, was paired with another subordinate and became dominant in this new pair, the inhibitory effect of 5-HT changed to an enhancing effect over the next 12 d of pairing. If, however, two dominant crayfish were paired and one became subordinate, the enhancing effect of 5-HT persisted in the new subordinate even after 38 d pairing. These different effects of serotonin result from the action of two or more molecular receptors for serotonin. A vertebrate 5-HT, agonist had no effect on social isolates but reversibly inhibited the response of LG in both dominant and subordinate crayfish. The inhibitory effects of the agonist developed approximately linearly over the first 12 d of pairing. A vertebrate 5-HT2 agonist persistently increased the response of LG in isolate crayfish and reversibly increased the response of the cell in dominant and subordinate crayfish. Finally, although neurons that might mediate these effects of superfused 5-HT are unknown, one pair of 5-HT-immunoreactive neurons appears to contact the LG axon and initial axon segment in each abdominal ganglion in its projection caudally from the thorax.
Figures
References
-
- Albers HE, Rawls S. Coordination of hamster lordosis and flank marking behavior: role of arginine vasopressin within the medial preoptic-anterior hypothalamus. Brain Res Bull. 1989;23:105–109. - PubMed
-
- Beall SP, Langley DJ, Edwards DH. Inhibition of escape tailflip in crayfish during backward walking and the defense posture. J Exp Biol. 1990;152:577–582. - PubMed
-
- Beltz BS, Kravitz EA. Aminergic and peptidergic neuromodulation in crustacea. J Exp Biol. 1986;124:115–141.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources