

Neuronal nicotinic receptor expression in sensory neurons of the rat trigeminal ganglion: demonstration of alpha3beta4, a novel subtype in the mammalian nervous system
- PMID: 8987817
- PMCID: PMC6579205
- DOI: 10.1523/JNEUROSCI.16-24-07892.1996
Neuronal nicotinic receptor expression in sensory neurons of the rat trigeminal ganglion: demonstration of alpha3beta4, a novel subtype in the mammalian nervous system
Abstract
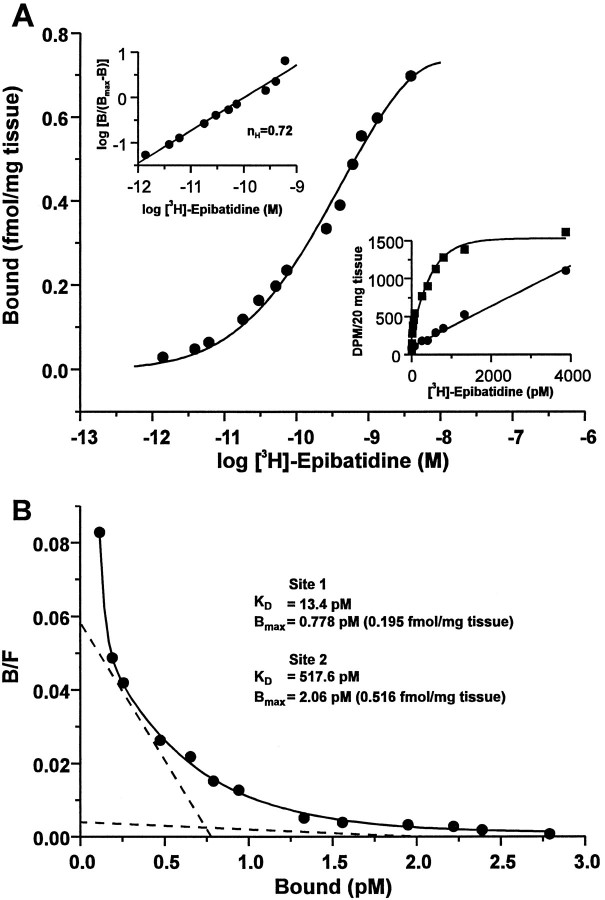
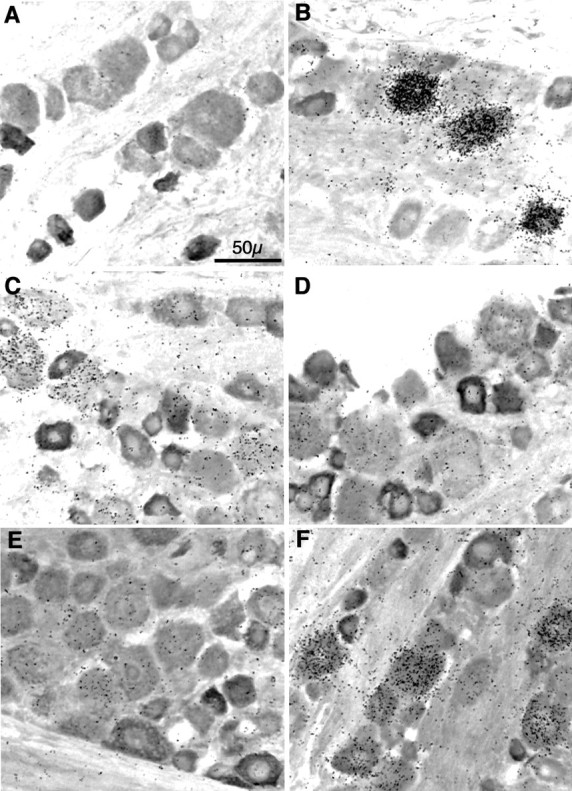
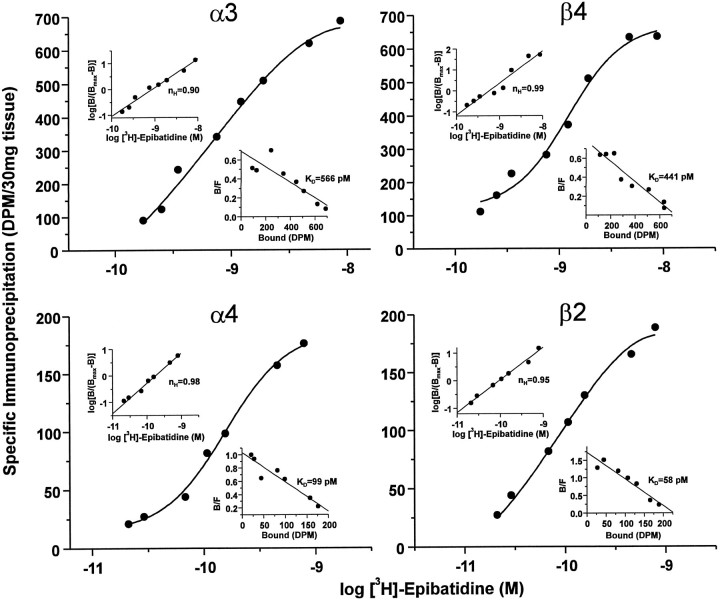
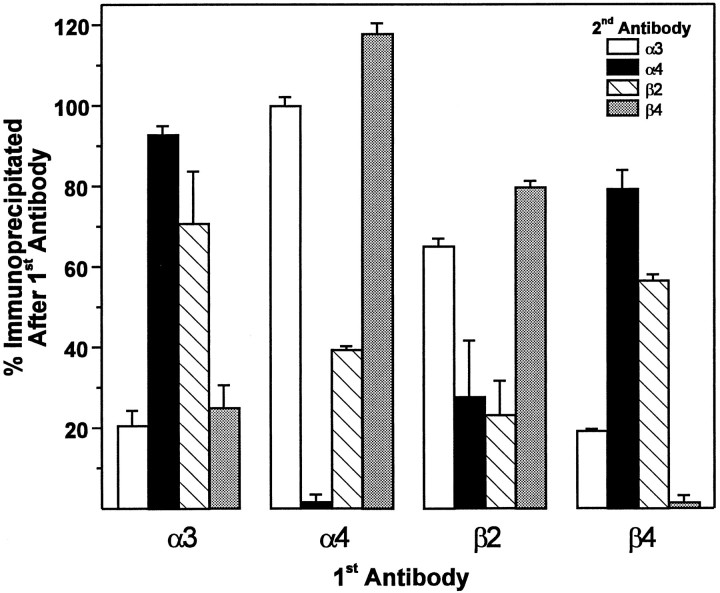
The identification of a family of neuronal nicotinic receptor subunit genes establishes the potential for multiple subtypes with diverse physiological functions. Virtually all of the high affinity nicotinic receptors measured to date in the rodent CNS are composed of alpha4 and beta2 subunits only. However, the demonstration of other subunit transcripts in a variety of central and peripheral nervous tissues suggests a greater degree of receptor subtype heterogeneity than so far has been elucidated. The purpose of the present studies was to determine at the mRNA and protein levels which neuronal nicotinic receptor subunits are expressed by sensory neurons of the rat trigeminal ganglion and in what combinations these gene products associate to form neuronal nicotinic receptor subtypes in this tissue. Radioreceptor binding analysis indicated that in the adult rat trigeminal ganglion there exist at least two nicotinic receptor binding sites with differing affinities for [3H]-epibatidine. In situ hybridization histochemical studies revealed the existence of mRNA encoding the alpha3, alpha4, alpha5, beta2, and beta4 subunits, but not the alpha2 subunit. Immunoprecipitation with subunit-specific antisera demonstrated that each of the subunits present in the ganglion at the mRNA level is a constituent of nicotinic receptors capable of binding 3H-epibatidine. Various applications of these approaches yielded strong evidence that, in addition to alpha4beta2, which is thought to be the predominant neuronal nicotinic receptor subtype in the rodent CNS, trigeminal sensory neurons express as the principal subtype alpha3beta4, which has not been demonstrated previously in mammalian nervous tissue.
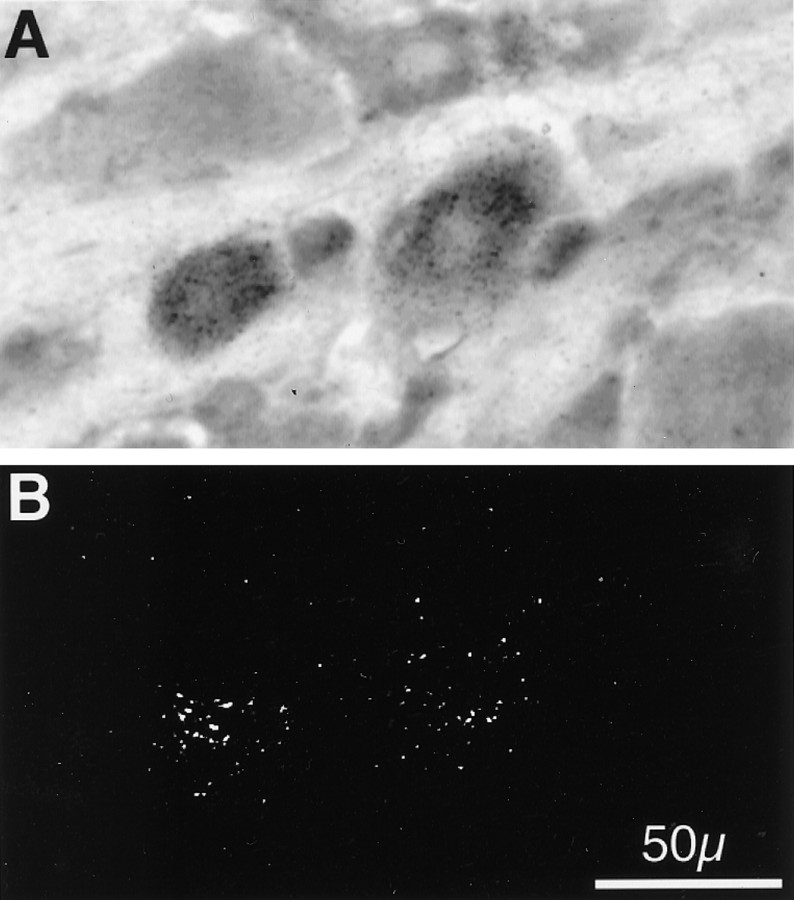
Figures

References
-
- Anand R, Conroy WG, Schoepfer R, Whiting P, Lindstrom J. Neuronal nicotinic acetylcholine receptors expressed in Xenopus oocytes have a pentameric quaternary structure. J Biol Chem. 1991;266:11192–11198. - PubMed
-
- Boulter J, Evans K, Goldman D, Martin G, Treco D, Heinemann S, Patrick J. Isolation of a cDNA clone coding for a possible neural nicotinic acetylcholine receptor α-subunit. Nature. 1986;319:368–374. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources