

Neuronal regulation of glutamate transporter subtype expression in astrocytes
- PMID: 8994048
- PMCID: PMC6573161
- DOI: 10.1523/JNEUROSCI.17-03-00932.1997
Neuronal regulation of glutamate transporter subtype expression in astrocytes
Abstract
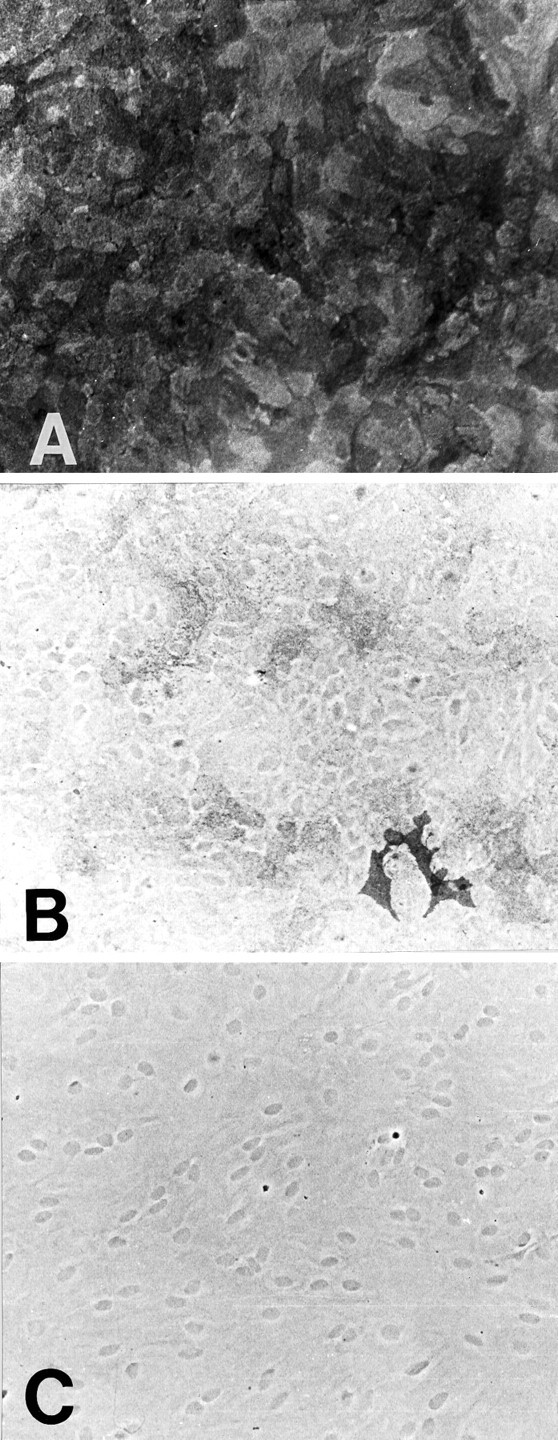
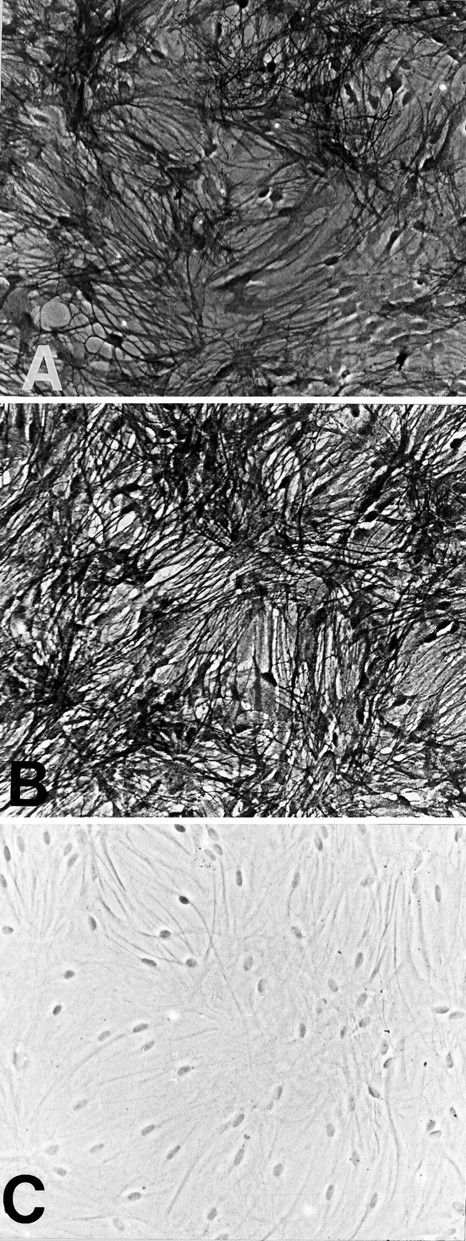
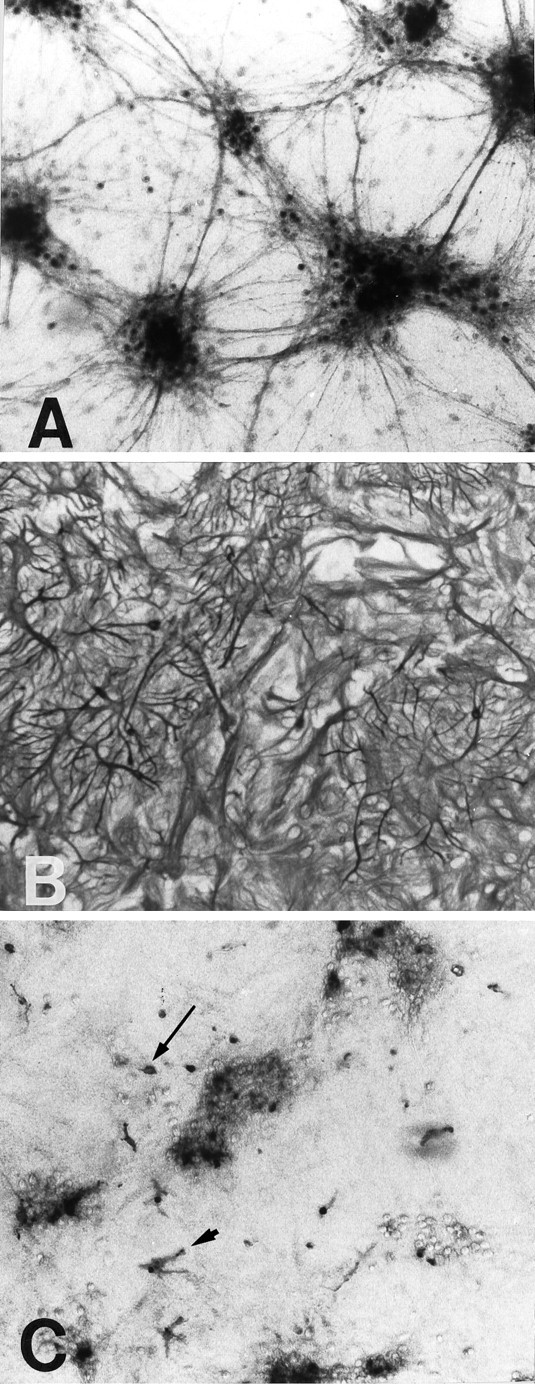
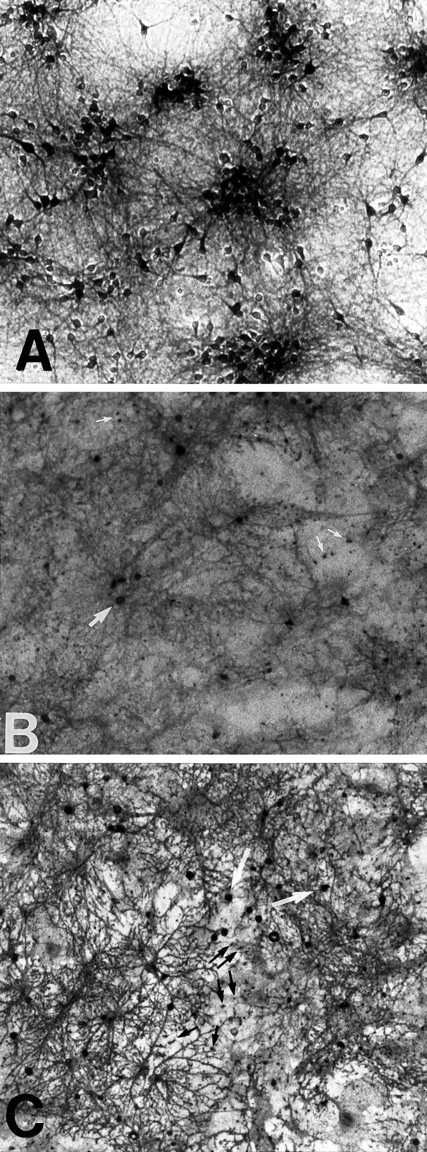
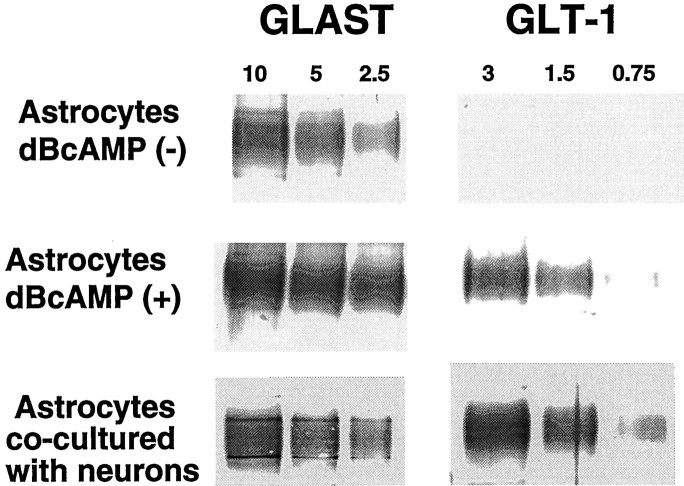
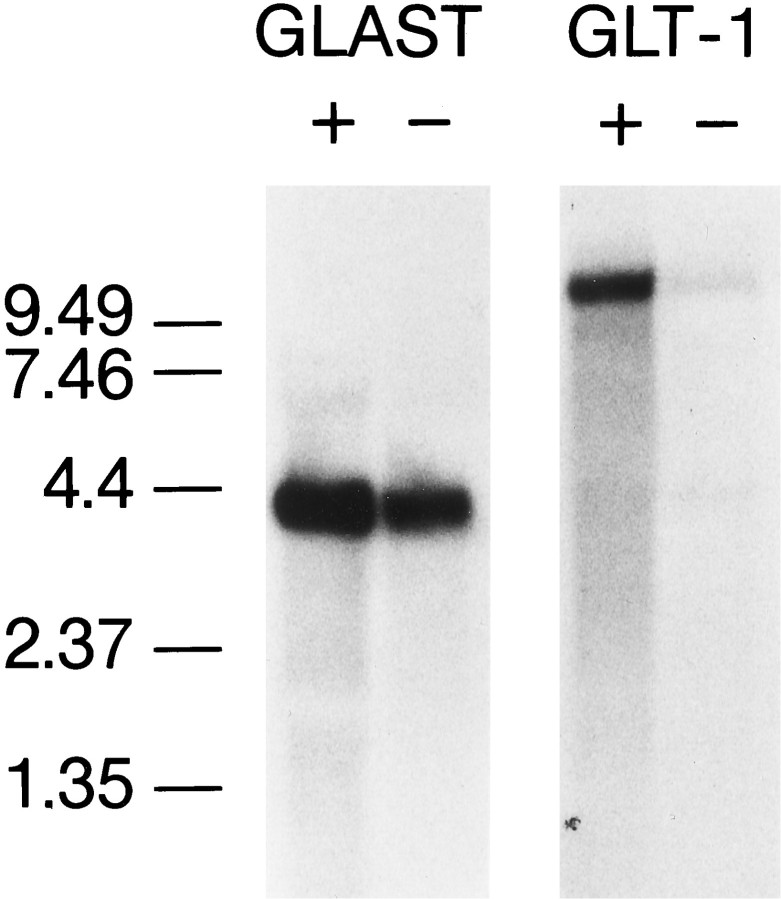
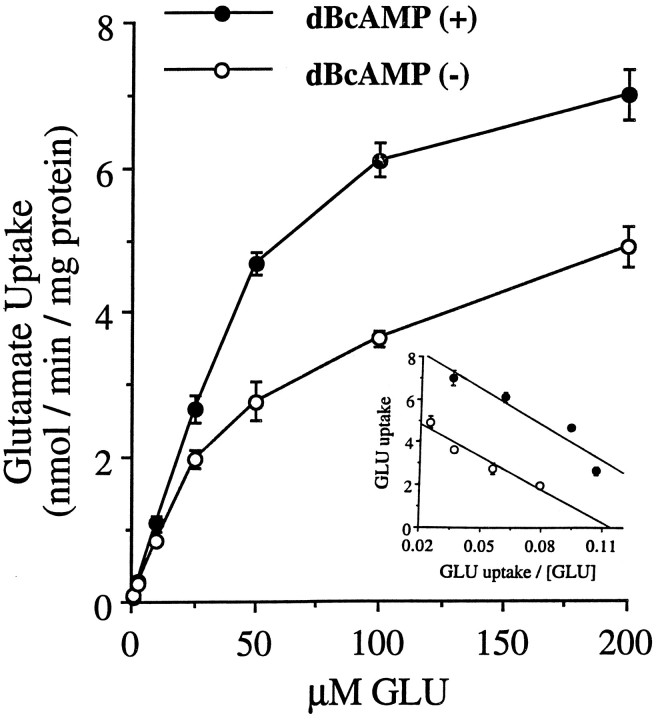
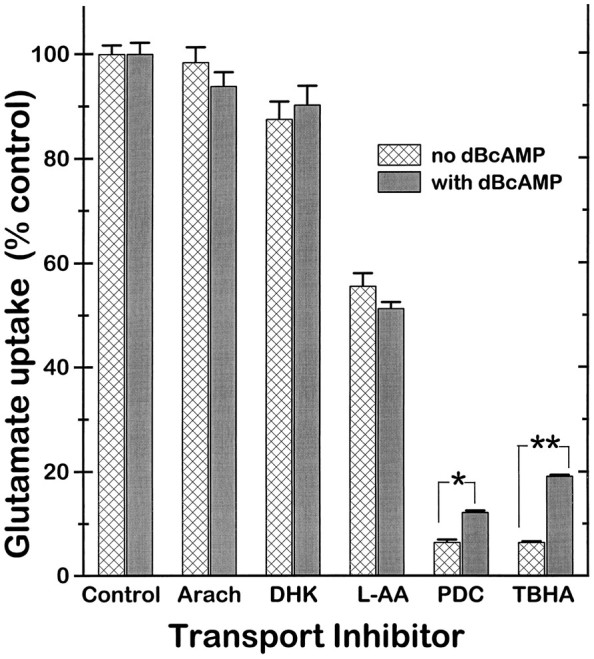
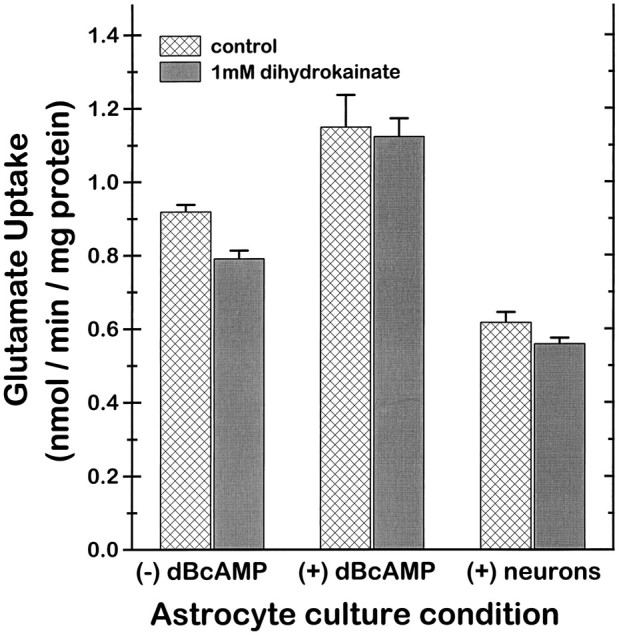
GLT-1, GLAST, and EAAC1 are high-affinity, Na(+)-dependent glutamate transporters identified in rat forebrain. The expression of these transporter subtypes was characterized in three preparations: undifferentiated rat cortical astrocyte cultures, astrocytes cocultured with cortical neurons, and astrocyte cultures differentiated with dibutyryl cyclic AMP (dBcAMP). The undifferentiated astrocyte monocultures expressed only the GLAST subtype. Astrocytes cocultured with neurons developed a stellate morphology and expressed both GLAST and GLT-1; neurons expressed only the EAAC1 transporter, and rare microglia in these cultures expressed GLT-1. Treatment of astrocyte cultures with dBcAMP induced expression of GLT-1 and increased expression of GLAST. These effects of dBcAMP on transporter expression were qualitatively similar to those resulting from coculture with neurons, but immunocytochemistry showed the pattern of transporter expression to be more complex in the coculture preparations. Compared with astrocytes expressing only GLAST, the dBcAMP-treated cultures expressing both GLAST and GLT-1 showed an increase in glutamate uptake Vmax, but no change in the glutamate K(m) and no increased sensitivity to inhibition by dihydrokainate. Pyrrolidine-2,4-dicarboxylic acid and threo-beta-hydroxyaspartic acid caused relatively less inhibition of transport in cultures expressing both GLAST and GLT-1, suggesting a weaker effect at GLT-1 than at GLAST. These studies show that astrocyte expression of glutamate transporter subtypes is influenced by neurons, and that dBcAMP can partially mimic this influence. Manipulation of transporter expression in astrocyte cultures may permit identification of factors regulating the expression and function of GLAST and GLT-1 in their native cell type.
Figures
References
-
- Bjørås M, Gjesdal O, Erickson JD, Torp R, Levy LM, Ottersen OP, Degree M, Storm-Mathisen J, Seeberg E, Danbolt NC. Cloning and expression of a neuronal rat brain glutamate transporter. Mol Brain Res. 1996;36:163–168. - PubMed
-
- Bristol LA, Rothstein JD. Glutamate transporter gene expression in amyotrophic lateral sclerosis motor cortex. Ann Neurol. 1996;39:676–679. - PubMed
-
- Cammer W. Glutamine synthetase in the central nervous system is not confined to astrocytes. J Neuroimmunol. 1990;26:173–178. - PubMed
-
- Castellano B, Gonzalez B, Jensen MB, Pedersen EB, Finsen BR, Zimmer J. A double staining technique for simultaneous demonstration of astrocytes and microglia in brain sections and astroglial cell cultures. J Histochem Cytochem. 1991;39:561–568. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources