

Song- and order-selective neurons in the songbird anterior forebrain and their emergence during vocal development
- PMID: 8994068
- PMCID: PMC6573158
- DOI: 10.1523/JNEUROSCI.17-03-01147.1997
Song- and order-selective neurons in the songbird anterior forebrain and their emergence during vocal development
Abstract
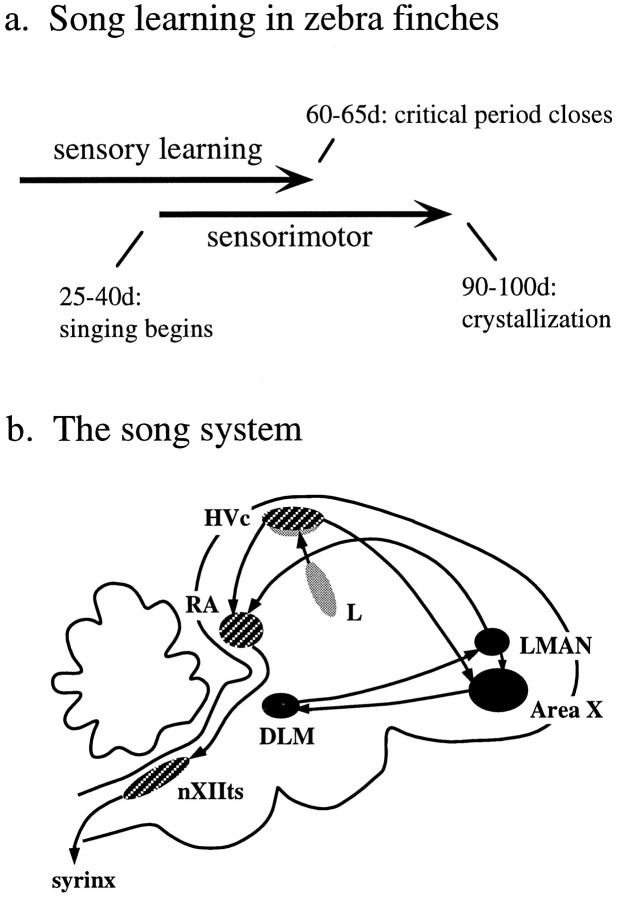
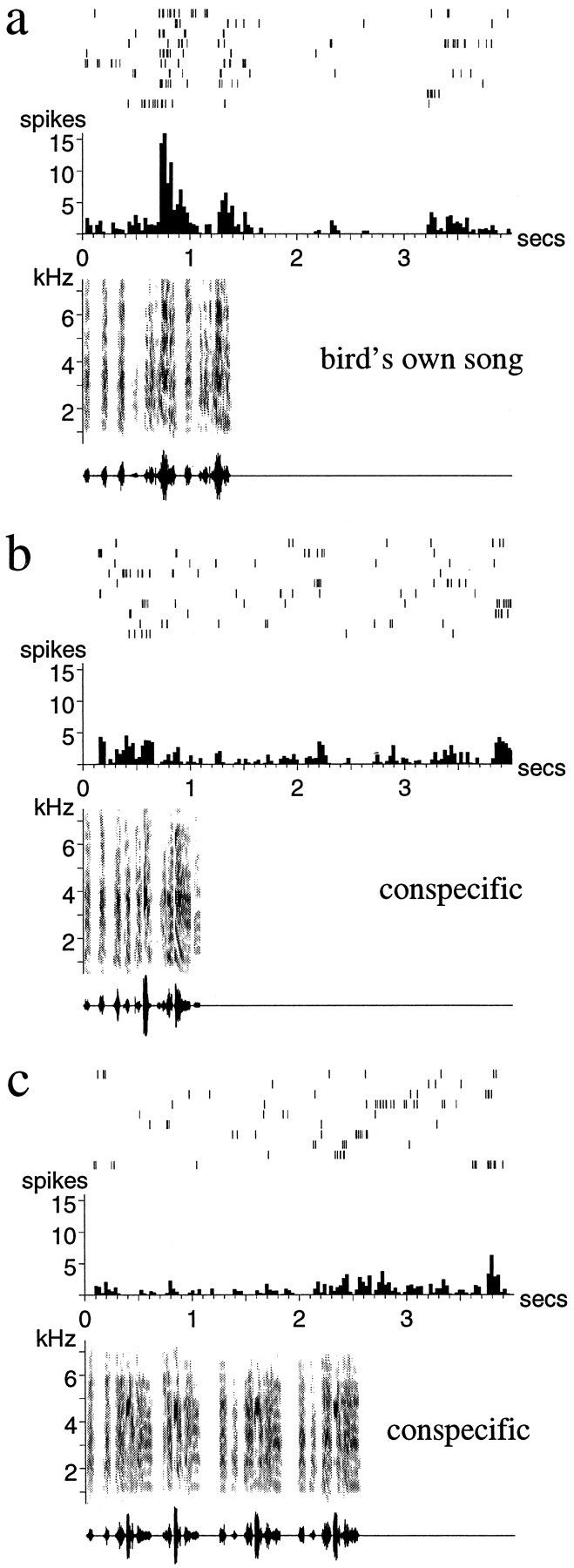
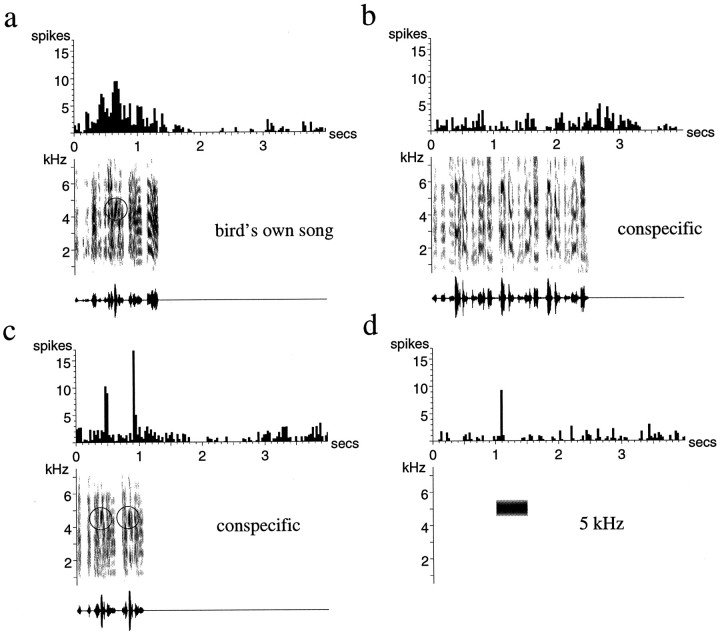
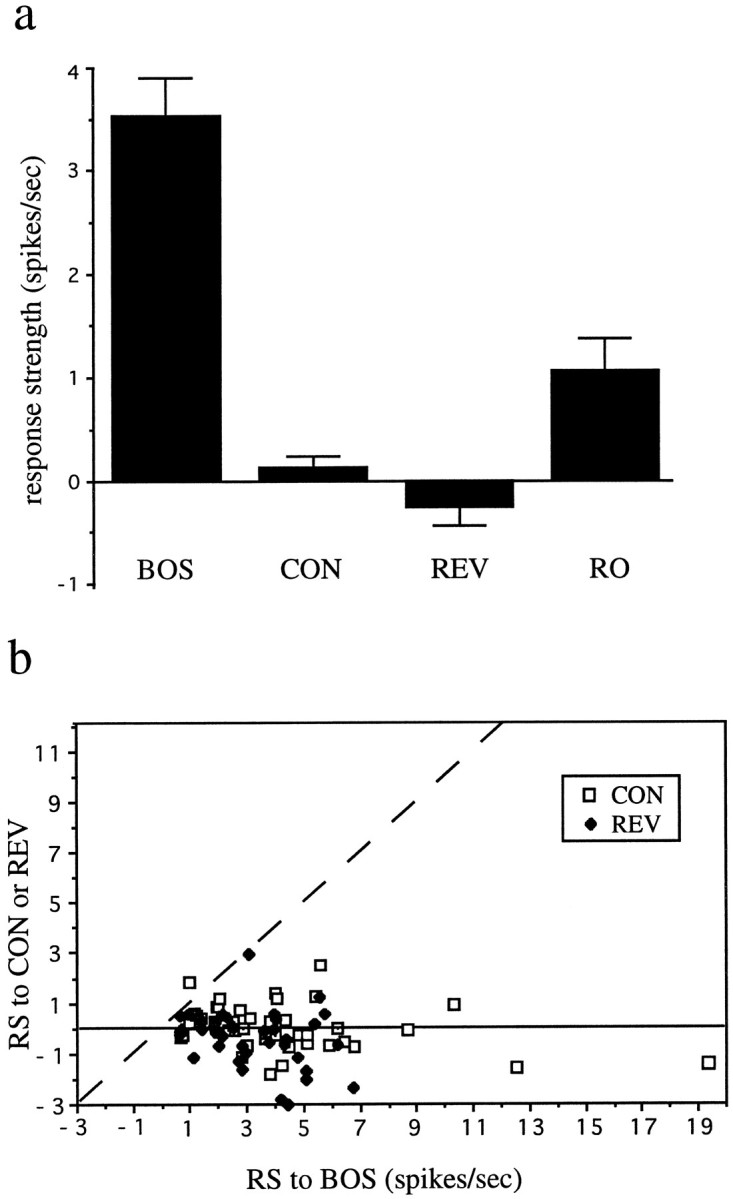
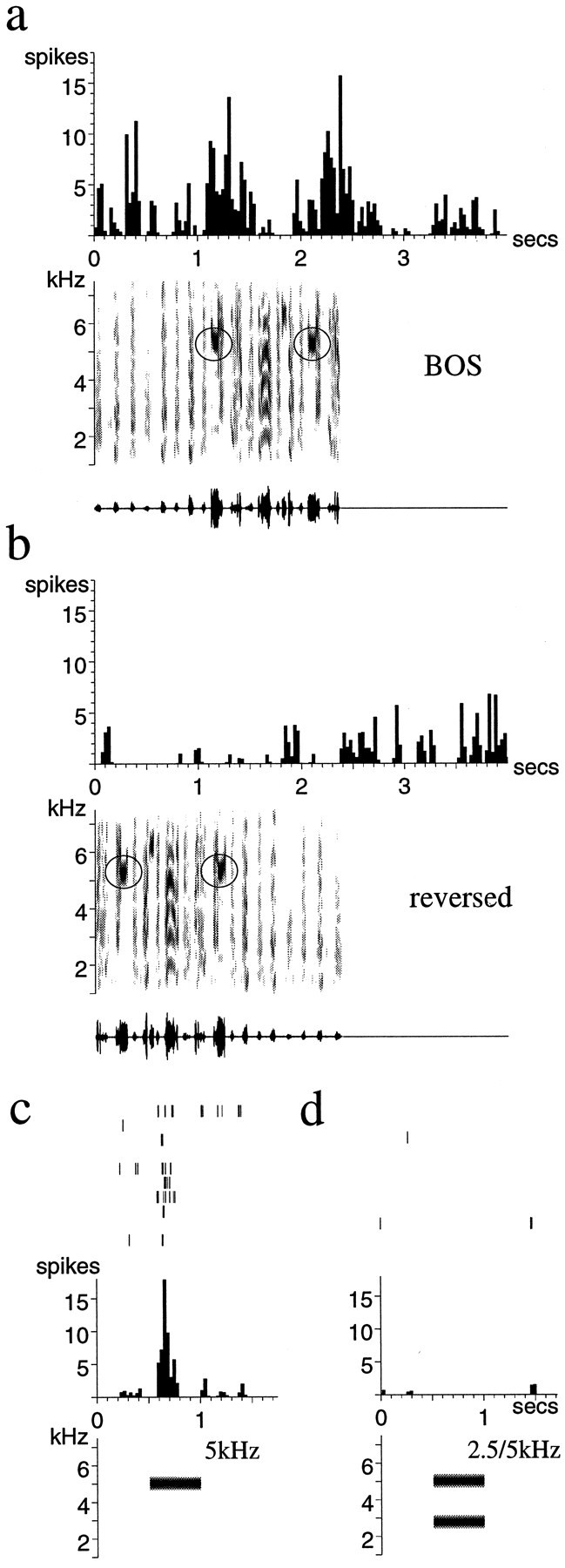
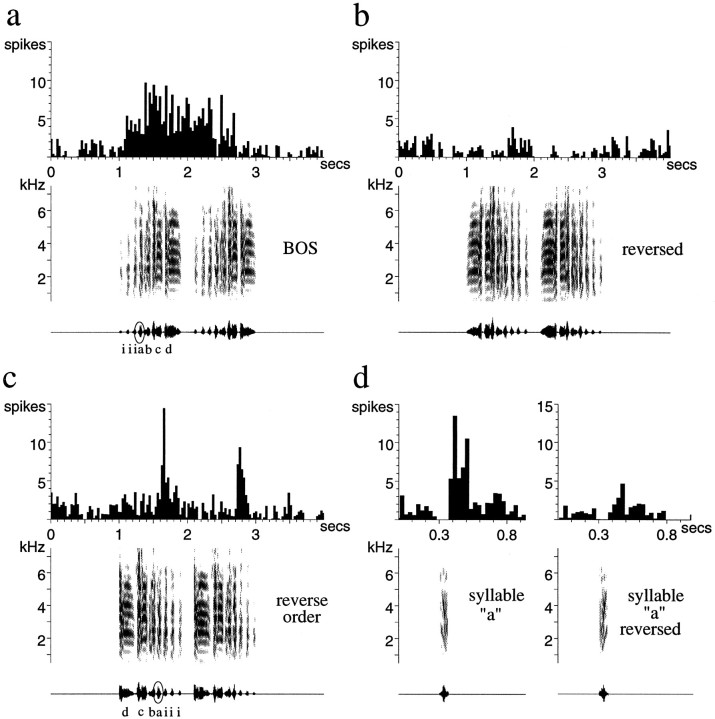
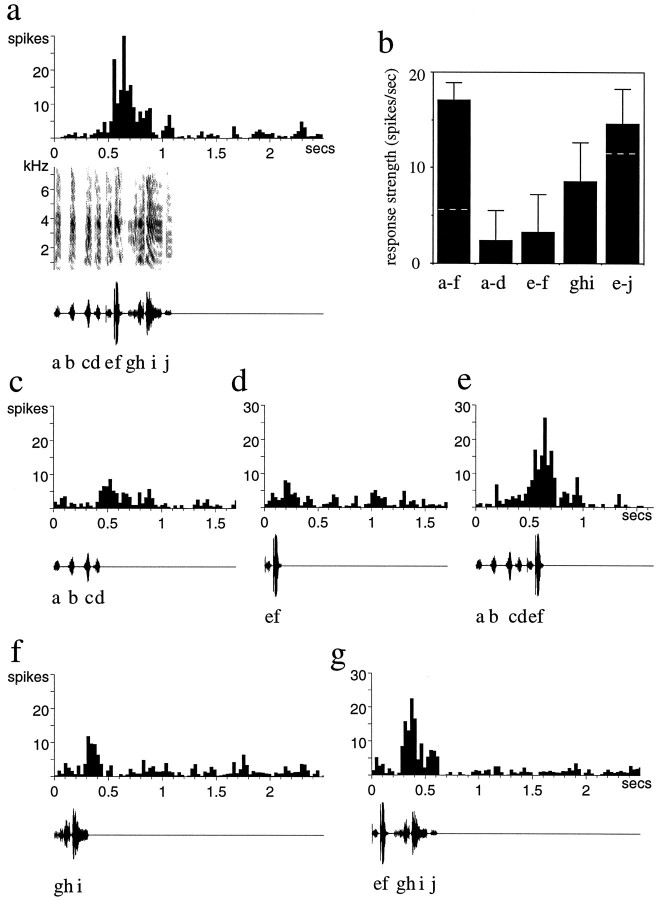
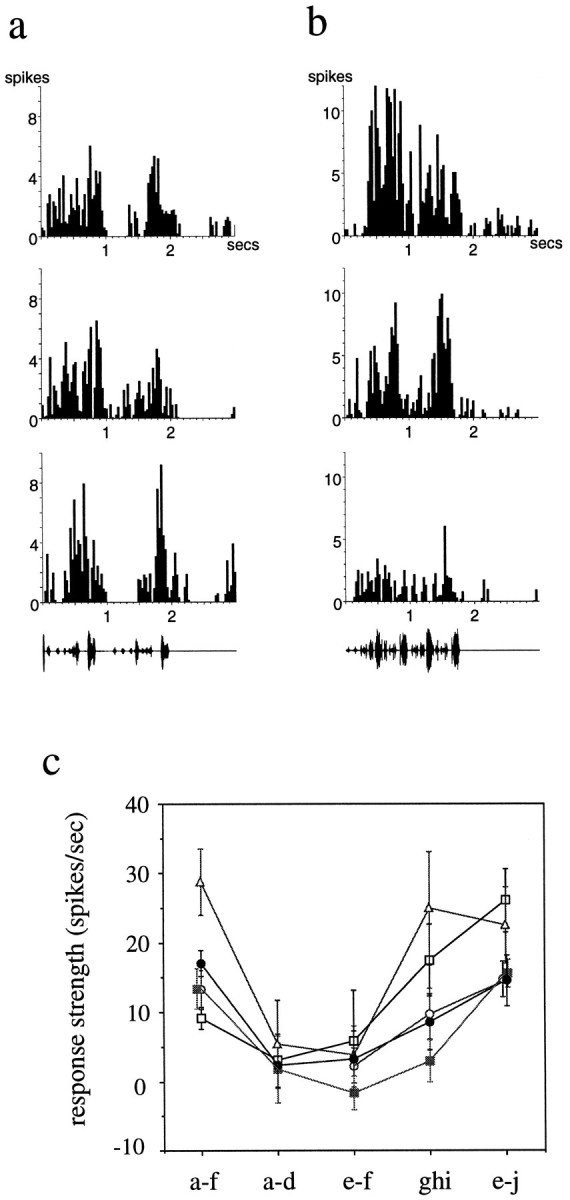
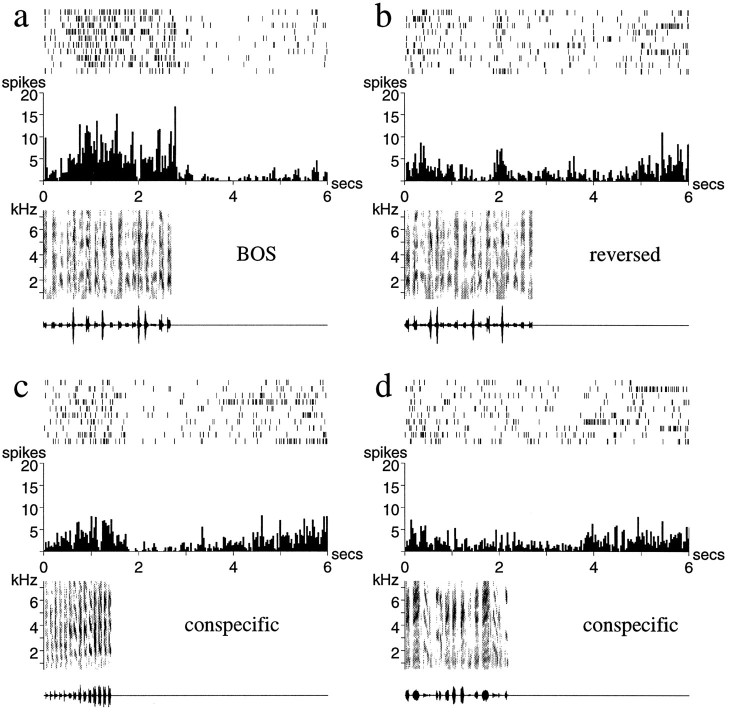
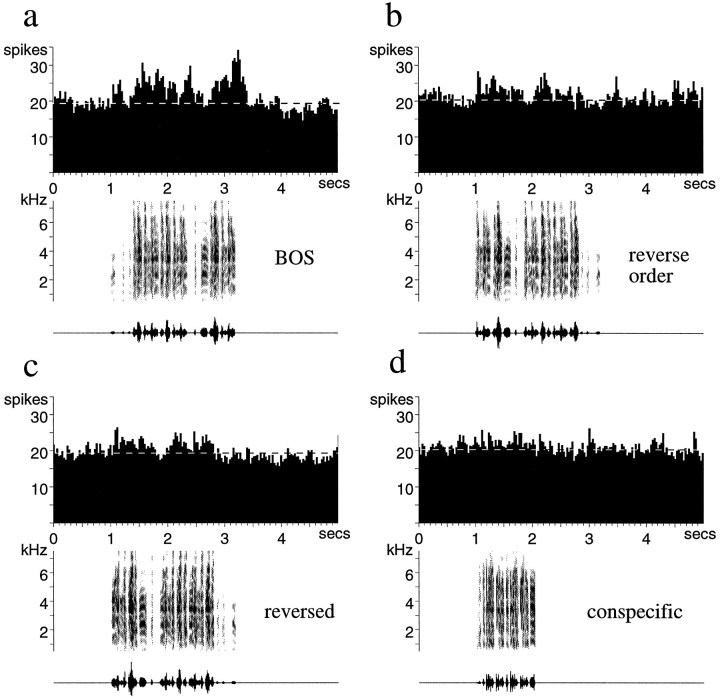
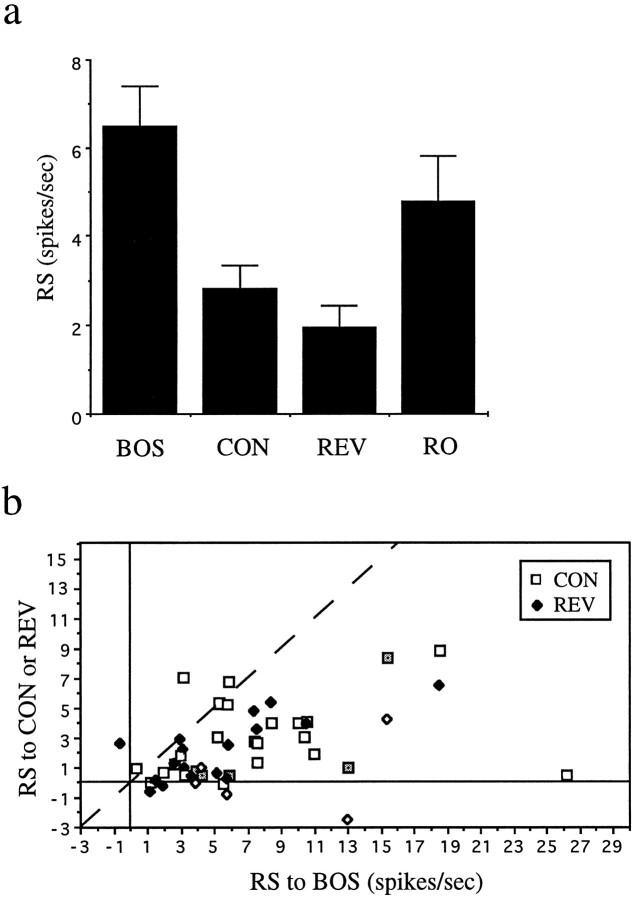
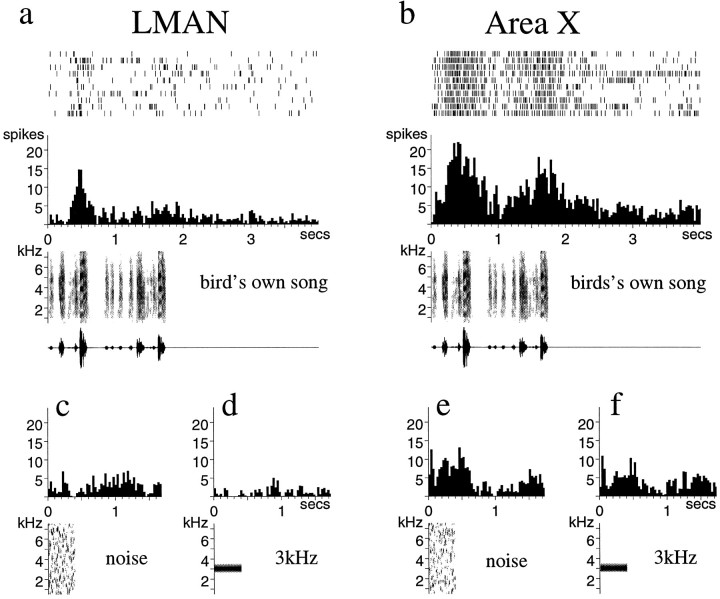
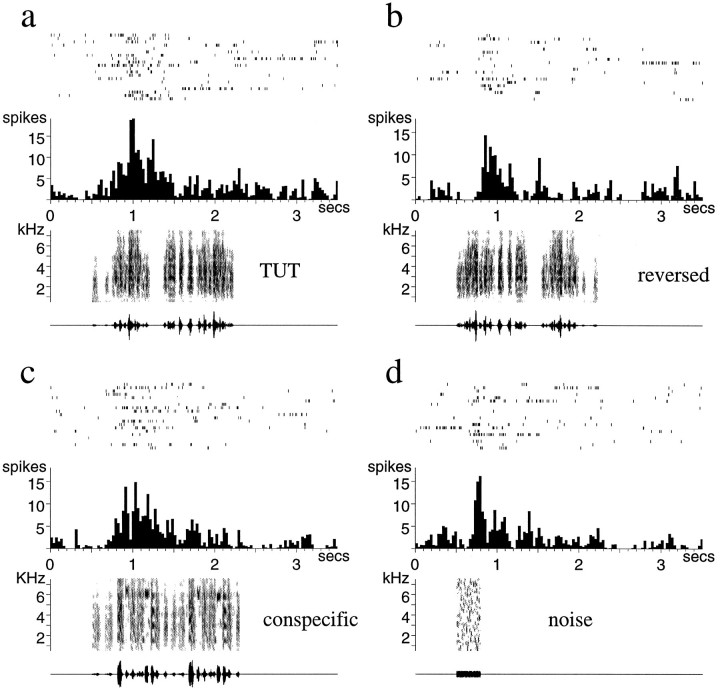
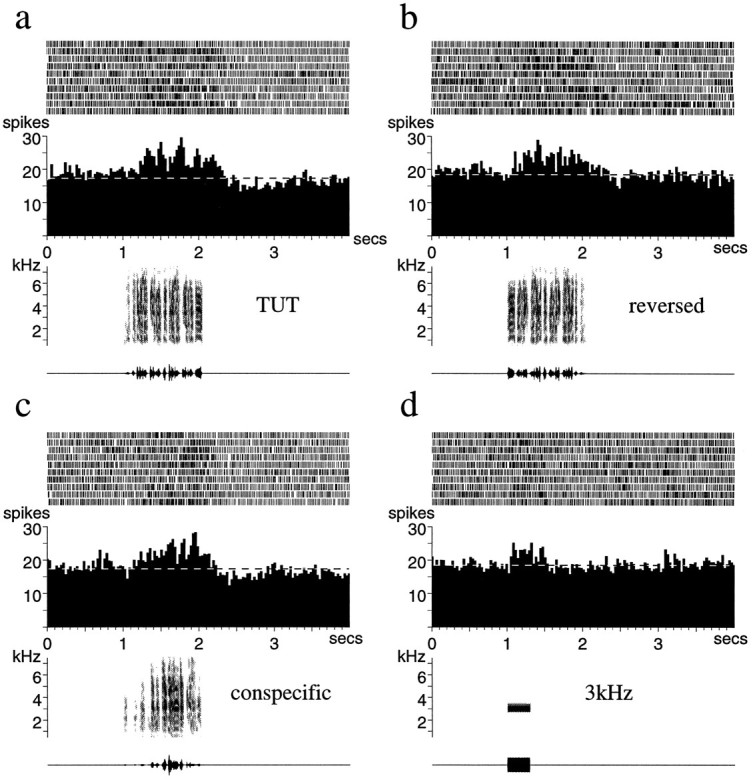
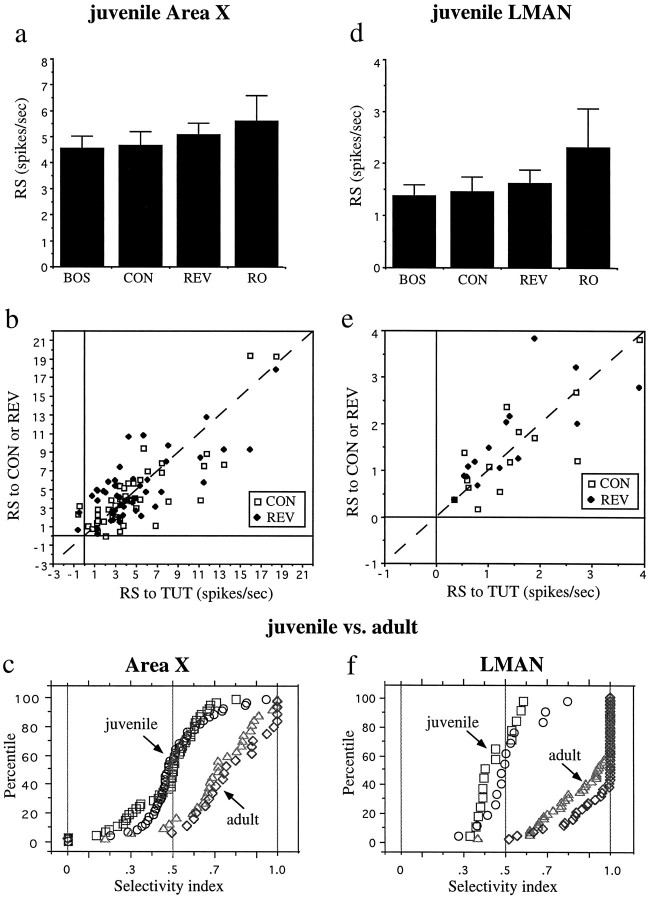
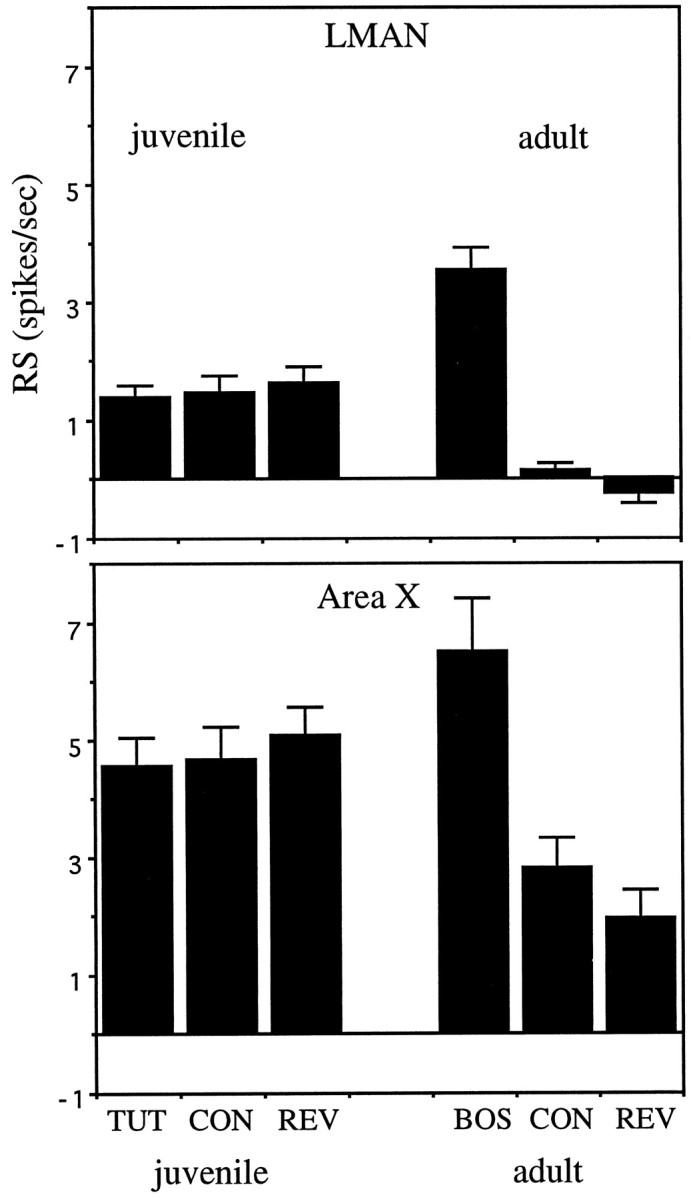
Auditory experience is critical for vocal learning in songbirds as in humans. Therefore, in a search for neural mechanisms for song learning and recognition, the auditory response properties of neurons in the anterior forebrain (AF) pathway of the songbird brain were investigated. This pathway plays an essential but poorly understood role during the period of song development when auditory feedback is most crucial. Single-unit recordings demonstrated that both the lateral magnocellular nucleus of the anterior neostriatum (LMAN) and Area X (X) contain auditory neurons in adult male finches. These neurons are strongly selective for both spectral and temporal properties of song; they respond more robustly to the bird's own song (BOS) than to songs of conspecific individuals, and they respond less well to the BOS if it is played in reverse. In addition, X neurons are more broadly responsive than LMAN neurons, suggesting that responses to song become progressively more refined along this pathway. Both X and LMAN of young male finches early in the process of song learning (30-45 d old) also contain song-responsive auditory neurons, but these juvenile neurons lack the song and order selectivity present in adult birds. The spectral and temporal selectivity of the adult AF auditory neurons therefore arises during development in neurons that are initially broadly song-responsive. These neurons provide one of the clearest examples of experience-dependent acquisition of complex stimulus selectivity. Moreover, the auditory properties of the AF circuit suggest that one of its functions may be to mediate the auditory learning and feedback so essential to song development.
Figures
References
-
- Aamodt SM, Kozlowski MR, Nordeen EJ, Nordeen KW. Distribution and developmental change in [3H]- MK-801 binding within zebra finch song nuclei. J Neurobiol. 1992;23:997–1005. - PubMed
-
- Arnold A. The effects of castration on song development in zebra finches, Poephila guttata. J Exp Zool. 1975;191:261–278. - PubMed
-
- Basham ME, Nordeen EJ, Nordeen KW. Blockade of NMDA receptors in the anterior forebrain impairs sensory acquisition in the zebra finch. Neurobiol Learn Mem. 1996;66:295–304. - PubMed
-
- Boehner J. Early acquisition of song in the zebra finch, Taeniopygia guttata. Anim Behav. 1990;39:369–374.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources