

Replacement of pre-T cell receptor signaling functions by the CD4 coreceptor
- PMID: 8996248
- PMCID: PMC2196103
- DOI: 10.1084/jem.185.1.121
Replacement of pre-T cell receptor signaling functions by the CD4 coreceptor
Abstract
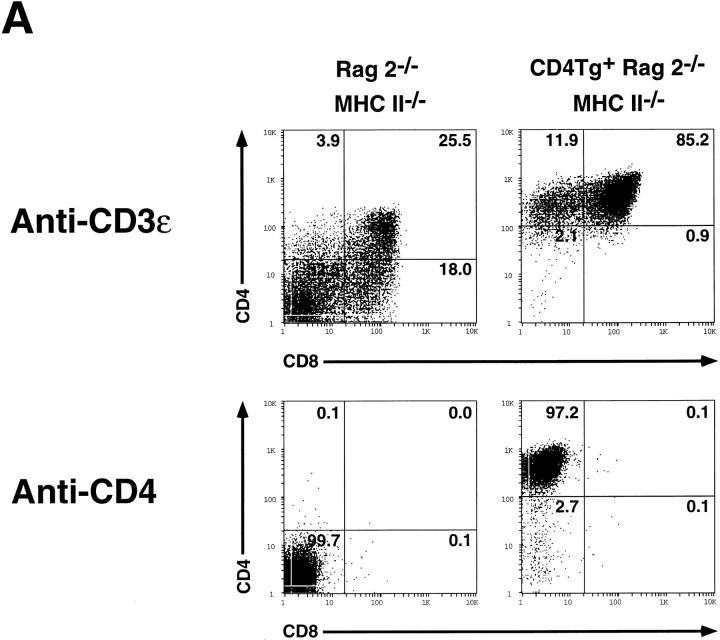
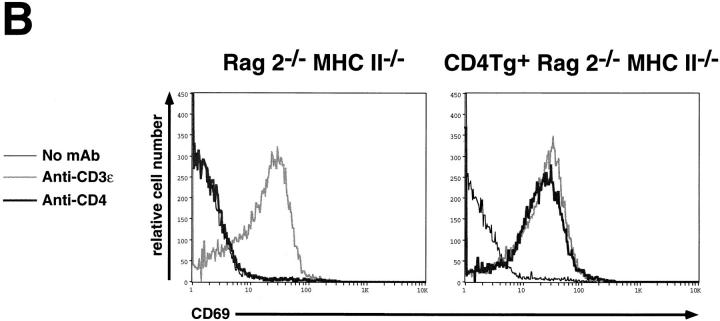
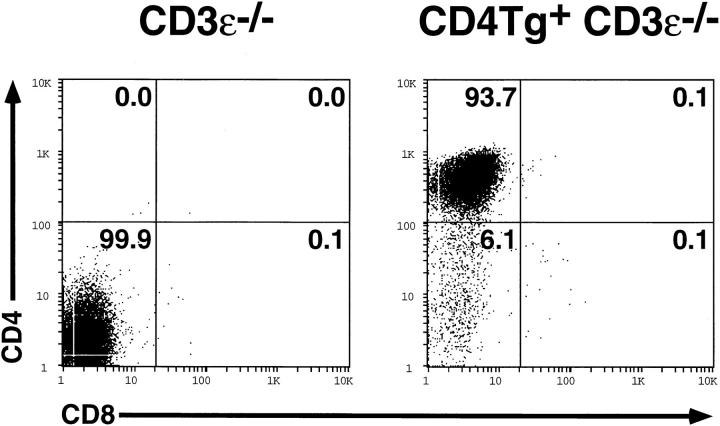
An important checkpoint in early thymocyte development ensures that only thymocytes with an in-frame T cell receptor for antigen beta (TCR-beta) gene rearrangement will continue to mature. Proper assembly of the TCR-beta chain into the pre-TCR complex delivers signals through the src-family protein tyrosine kinase p56lck that stimulate thymocyte proliferation and differentiation to the CD4+CD8+ stage. However, the biochemical mechanisms governing p56lck activation remain poorly understood. In more mature thymocytes, p56lck is associated with the cytoplasmic domain of the TCR coreceptors CD4 and CD8, and cross-linking of CD4 leads to p56lck activation. To study the effect of synchronously inducing p56lck activation in immature CD4-CD8- thymocytes, we generated mice expressing a CD4 transgene in Rag2-/- thymocytes. Remarkably, without further experimental manipulation, the CD4 transgene drives maturation of Rag2-/- thymocytes in vivo. We show that this process is dependent upon the ability of the CD4 transgene to bind Lck and on the expression of MHC class II molecules. Together these results indicate that binding of MHC class II molecules to CD4 can deliver a biologically relevant, Lck-dependent activation signal to thymocytes in the absence of the TCR-alpha or -beta chain.
Figures
References
-
- Perlmutter RM, Levin SD, Appleby MW, Anderson S J, Alberola-Ila J. Regulation of lymphocyte function by protein phosphorylation. Ann Rev Immunol. 1993;11:451–499. - PubMed
-
- Jenkinson EJ, Owen JJ. T-cell differentiation in thymus organ cultures. Semin Immunol. 1990;2:51–58. - PubMed
-
- Saint-Ruf C, Ungewiss K, Groettrup M, Bruno L, Fehling HJ, von Boehmer H. Analysis and expression of a cloned pre-T cell receptor gene. Science (Wash DC) 1994;266:1208–1212. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
