

The N-terminal domain of a K+ channel beta subunit increases the rate of C-type inactivation from the cytoplasmic side of the channel
- PMID: 9005448
- PMCID: PMC26366
- DOI: 10.1073/pnas.93.26.15119
The N-terminal domain of a K+ channel beta subunit increases the rate of C-type inactivation from the cytoplasmic side of the channel
Abstract
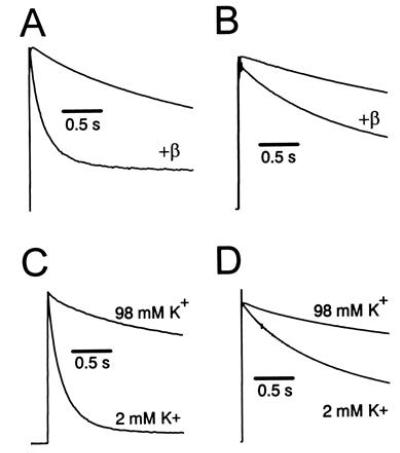
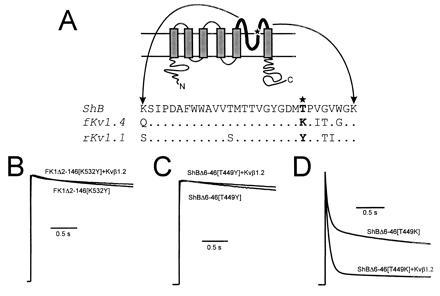
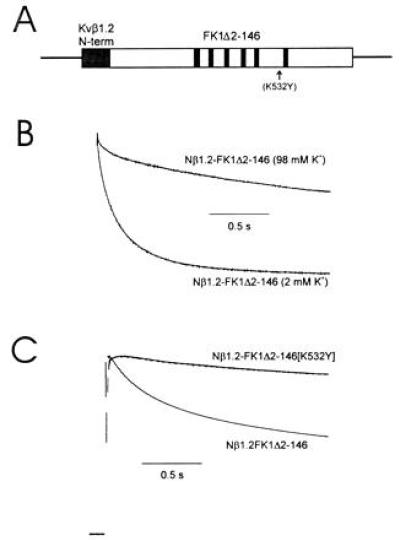
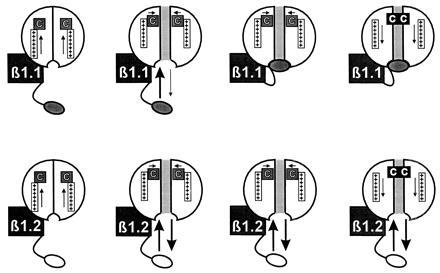
Voltage-gated K+ channels are complexes of membrane-bound, ion-conducting alpha and cytoplasmic ancillary (beta) subunits. The primary physiologic effect of coexpression of alpha and beta subunits is to increase the intrinsic rate of inactivation of the alpha subunit. For one beta subunit, Kv beta 1.1, inactivation is enhanced through an N-type mechanism. A second beta subunit, Kv beta 1.2, has been shown to increase inactivation, but through a distinct mechanism. Here we show that the degree of enhancement of Kv beta 1.2 inactivation is dependent on the amino acid composition in the pore mouth of the alpha subunit and the concentration of extracellular K+. Experimental conditions that promote C-type inactivation also enhance the stimulation of inactivation by Kv beta 1.2, showing that this beta subunit directly stimulates C-type inactivation. Chimeric constructs containing just the nonconserved N-terminal region of Kv beta 1.2 fused with an alpha subunit behave in a similar fashion to coexpressed Kv beta 1.2 and alpha subunit. This shows that it is the N-terminal domain of Kv beta 1.2 that mediates the increase in C-type inactivation from the cytoplasmic side of the pore. We propose a model whereby the N terminus of Kv beta 1.2 acts as a weakly binding "ball" domain that associates with the intracellular vestibule of the alpha subunit to effect a conformational change leading to enhancement of C-type inactivation.
Figures
References
-
- Chandy K G, Gutman G A. In: Handbook of Receptors and Channels: Ligand and Voltage-Gated Ion Channels. North R A, editor. Boca Raton, FL: CRC; 1995. pp. 1–71.
-
- Rettig J, Heinemann S H, Wunder F, Lorra C, Parcej D N, Dolly J O, Pongs O. Nature (London) 1994;369:289–294. - PubMed
-
- Morales M J, Castellino R C, Crews A L, Rasmusson R L, Strauss H C. J Biol Chem. 1995;270:6272–6277. - PubMed
-
- Majumder K, De Biasi M, Wang Z, Wible B A. FEBS Lett. 1995;361:13–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
