

Ligand-gated ion channel subunit partnerships: GABAA receptor alpha6 subunit gene inactivation inhibits delta subunit expression
- PMID: 9006978
- PMCID: PMC6793744
- DOI: 10.1523/JNEUROSCI.17-04-01350.1997
Ligand-gated ion channel subunit partnerships: GABAA receptor alpha6 subunit gene inactivation inhibits delta subunit expression
Abstract
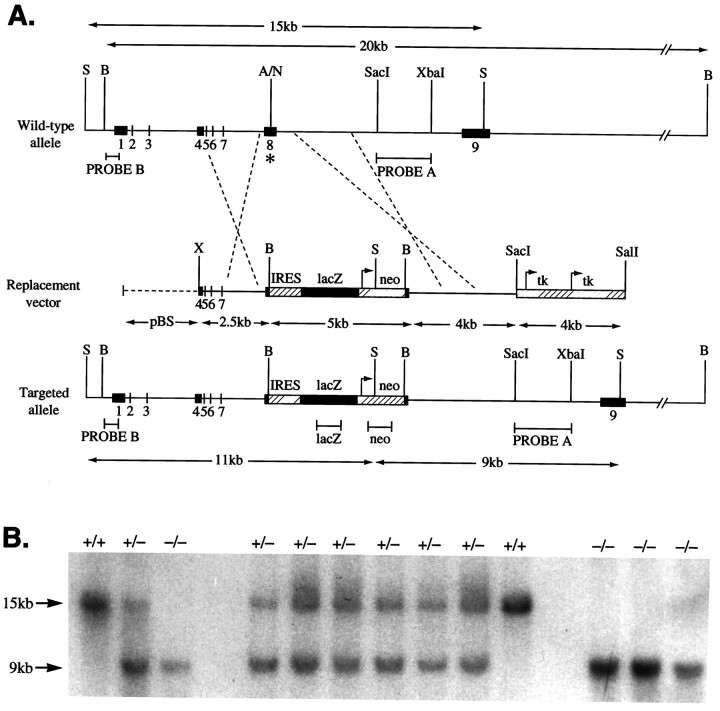
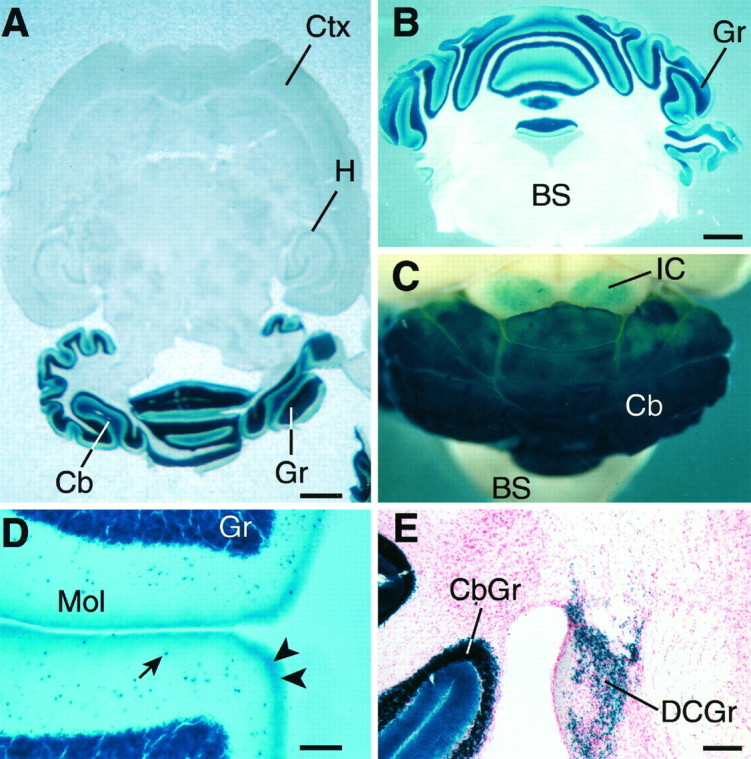
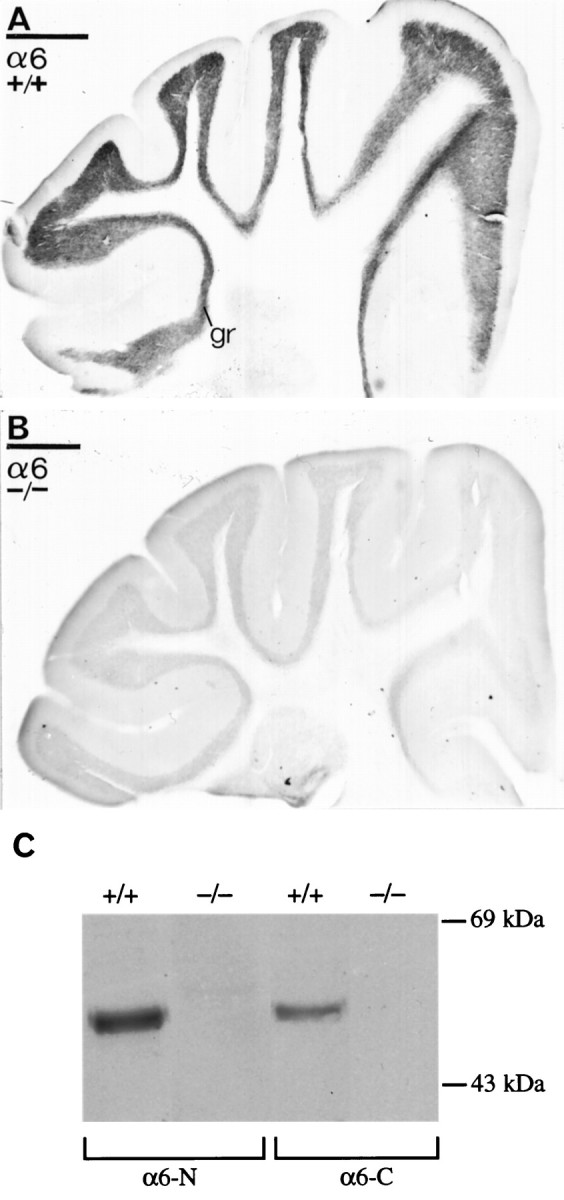
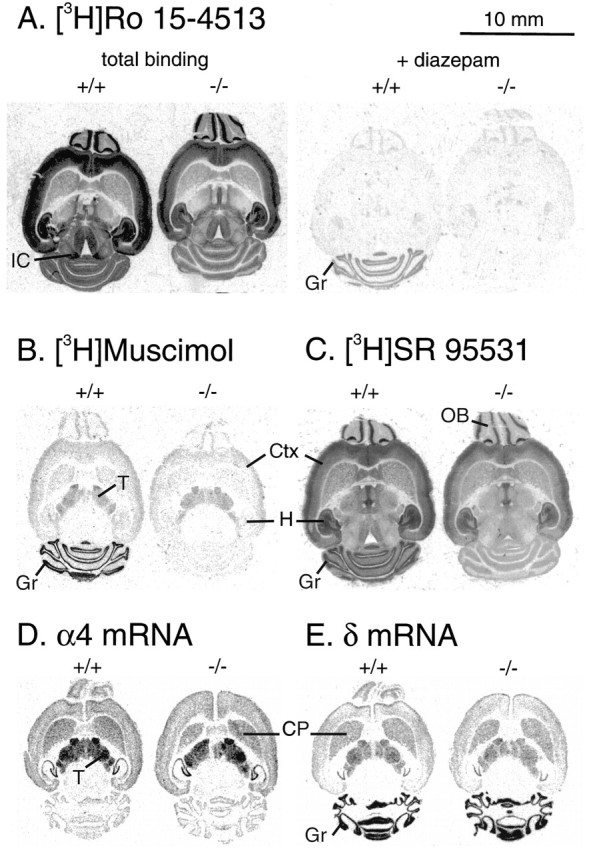
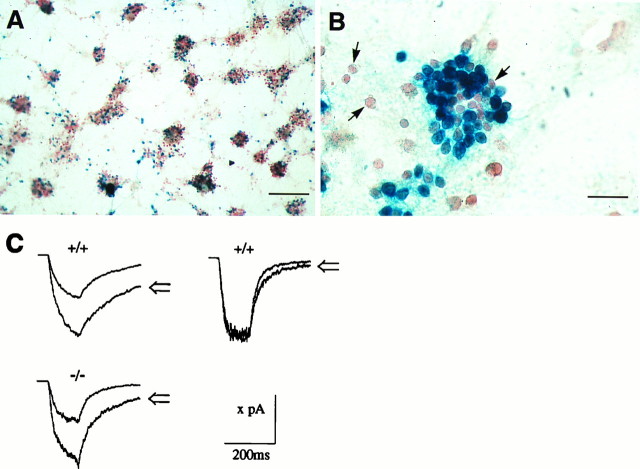
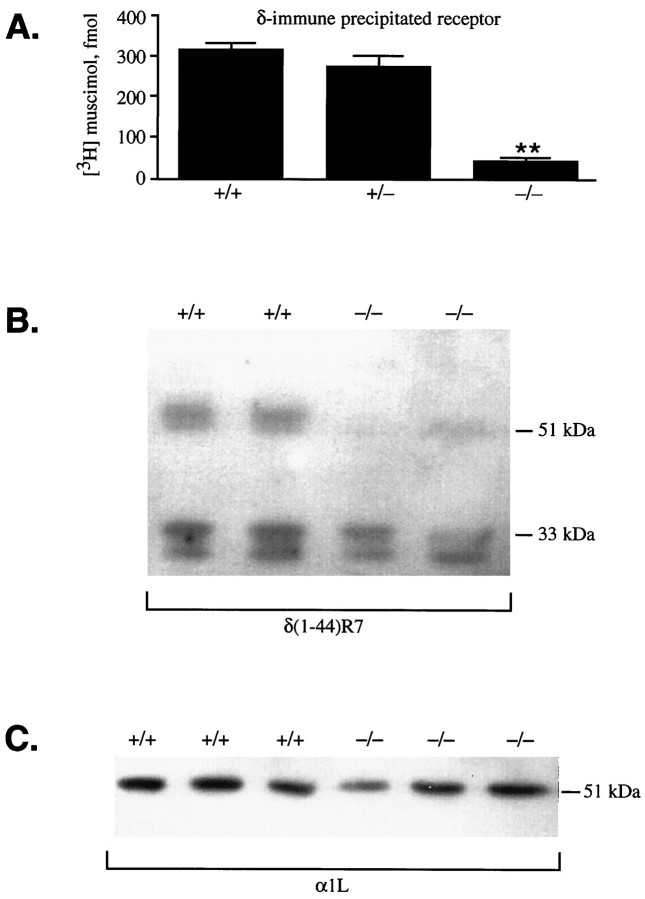
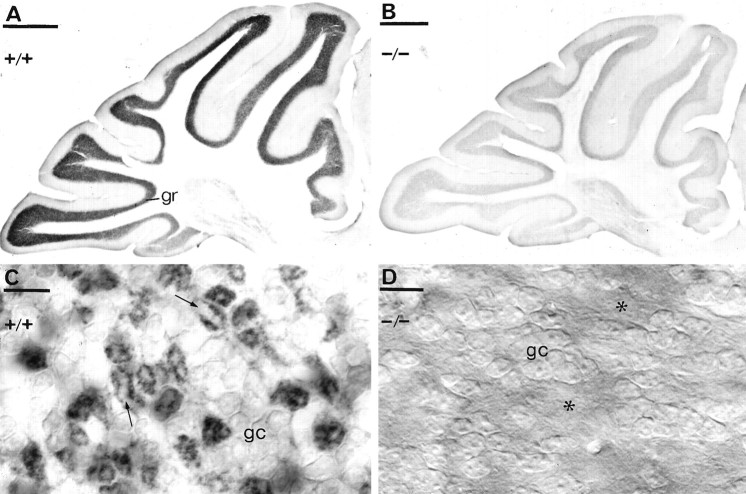
Cerebellar granule cells express six GABAA receptor subunits abundantly (alpha1, alpha6, beta2, beta3, gamma2, and delta) and assemble various pentameric receptor subtypes with unknown subunit compositions; however, the rules guiding receptor subunit assembly are unclear. Here, removal of intact alpha6 protein from cerebellar granule cells allowed perturbations in other subunit levels to be studied. Exon 8 of the mouse alpha6 subunit gene was disrupted by homologous recombination. In alpha6 -/- granule cells, the delta subunit was selectively degraded as seen by immunoprecipitation, immunocytochemistry, and immunoblot analysis with delta subunit-specific antibodies. The delta subunit mRNA was present at wild-type levels in the mutant granule cells, indicating a post-translational loss of the delta subunit. These results provide genetic evidence for a specific association between the alpha6 and delta subunits. Because in alpha6 -/- neurons the remaining alpha1, beta2/3, and gamma2 subunits cannot rescue the delta subunit, certain potential subunit combinations may not be found in wild-type cells.
Figures
References
-
- Bahn S, Harvey RJ, Darlison MG, Wisden W. Conservation of γ aminobutyric acid type A receptor α6 subunit gene expression in cerebellar granule cells. J Neurochem. 1996;66:1810–1818. - PubMed
-
- Benke D, Mertens S, Trzecia A, Gillessen D, Möhler H. Identification and immunohistochemical mapping of GABAA receptor subtypes containing the δ subunit in rat brain. FEBS Lett. 1991;283:145–149. - PubMed
-
- Bonnerot C, Nicolas J-F. Application of LacZ gene fusions to postimplantation development. Methods Enzymol. 1993;225:451–469. - PubMed
-
- Caruncho HJ, Costa E. Double-immunolabelling study of GABAA receptor subunits in label-fracture replicas of cultured rat cerebellar granule cells. Receptors Channels. 1994;2:143–153. - PubMed
-
- Caruncho HJ, Puia G, Möhler H, Costa E. The density and distribution of six GABAA receptor subunits in primary cultures of rat cerebellar granule cells. Neuroscience. 1995;67:583–593. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases