

A pacemaker current in dye-coupled hilar interneurons contributes to the generation of giant GABAergic potentials in developing hippocampus
- PMID: 9006985
- PMCID: PMC6793743
- DOI: 10.1523/JNEUROSCI.17-04-01435.1997
A pacemaker current in dye-coupled hilar interneurons contributes to the generation of giant GABAergic potentials in developing hippocampus
Abstract
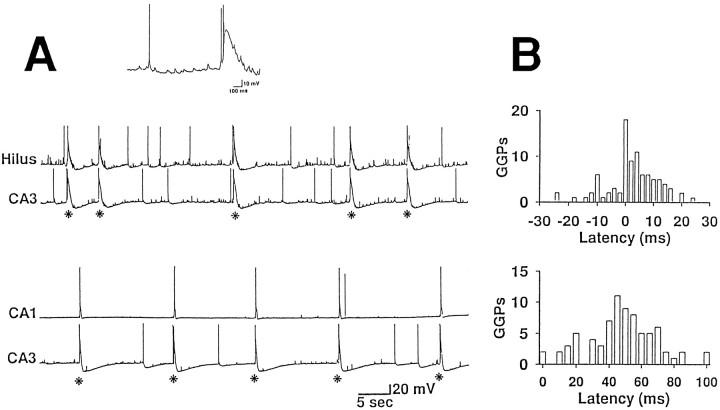
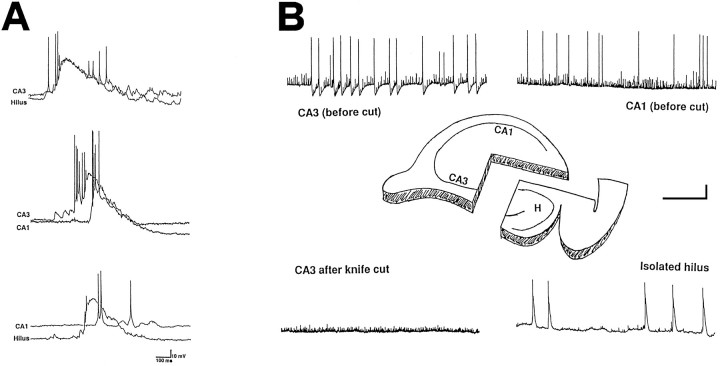
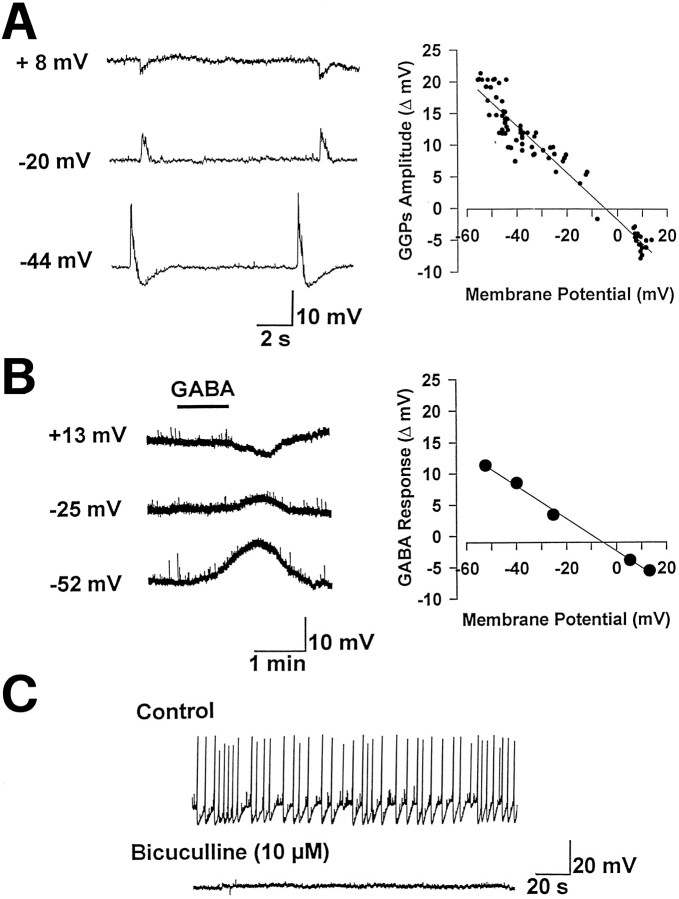
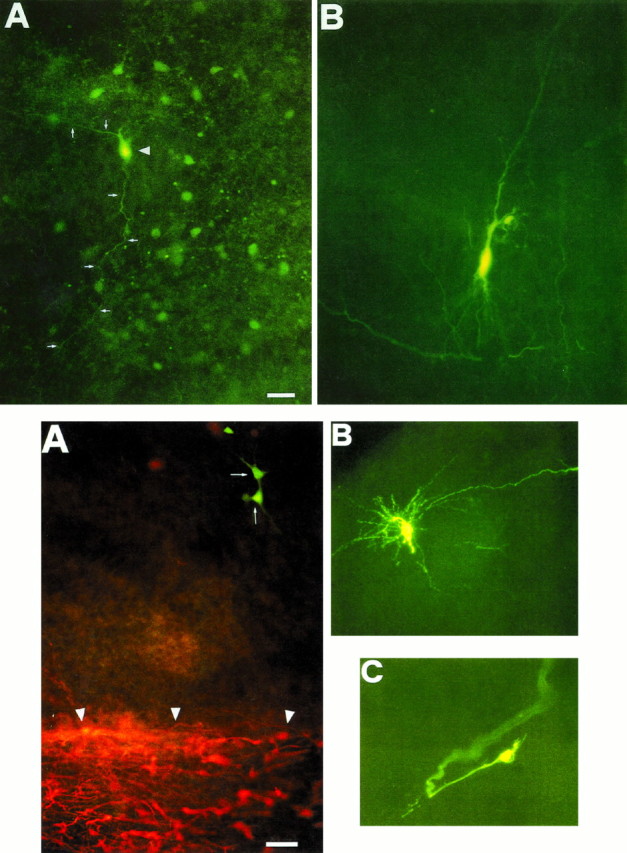
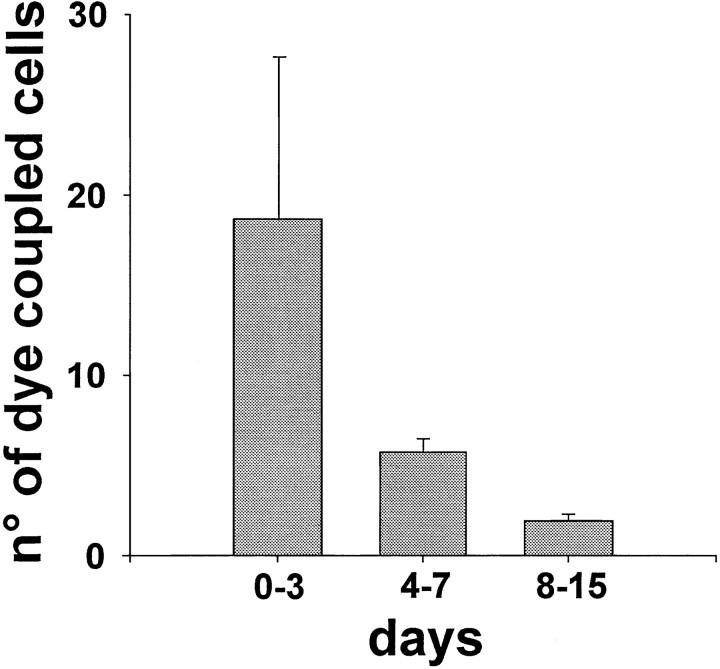
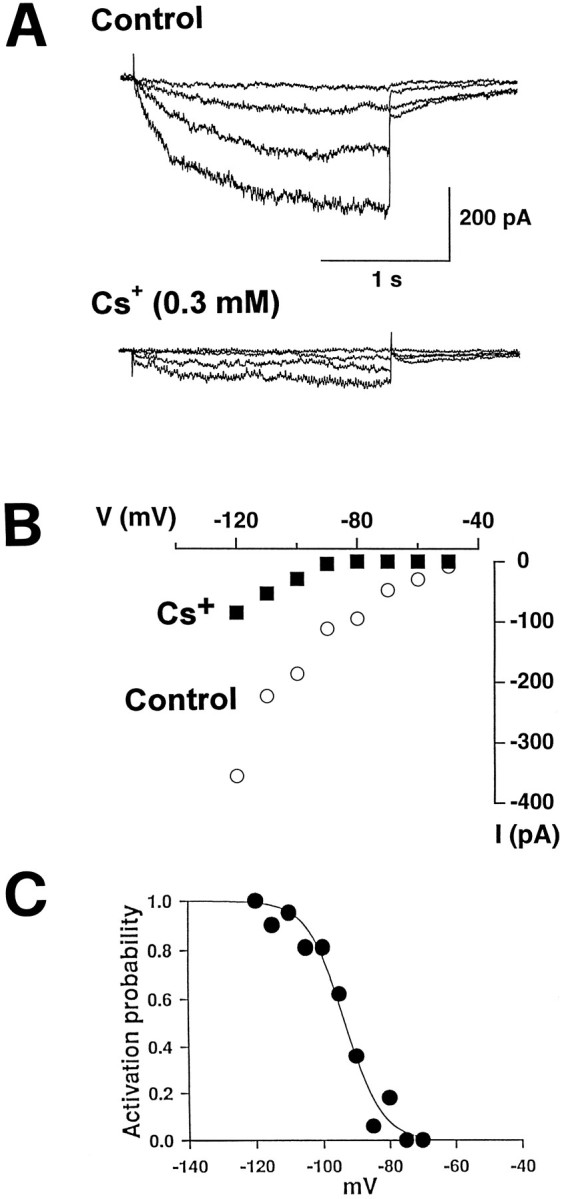
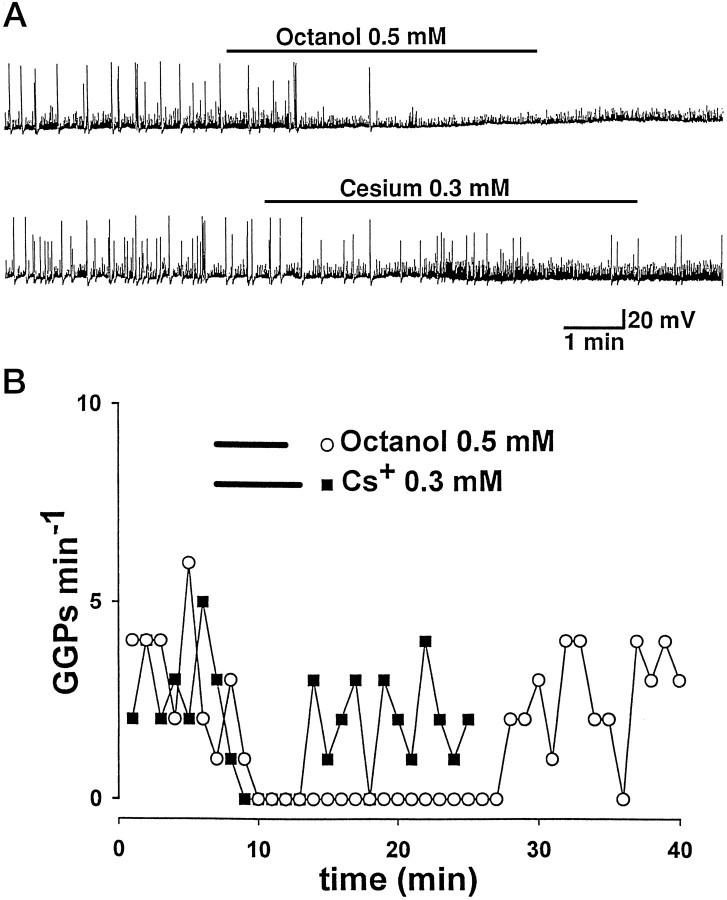
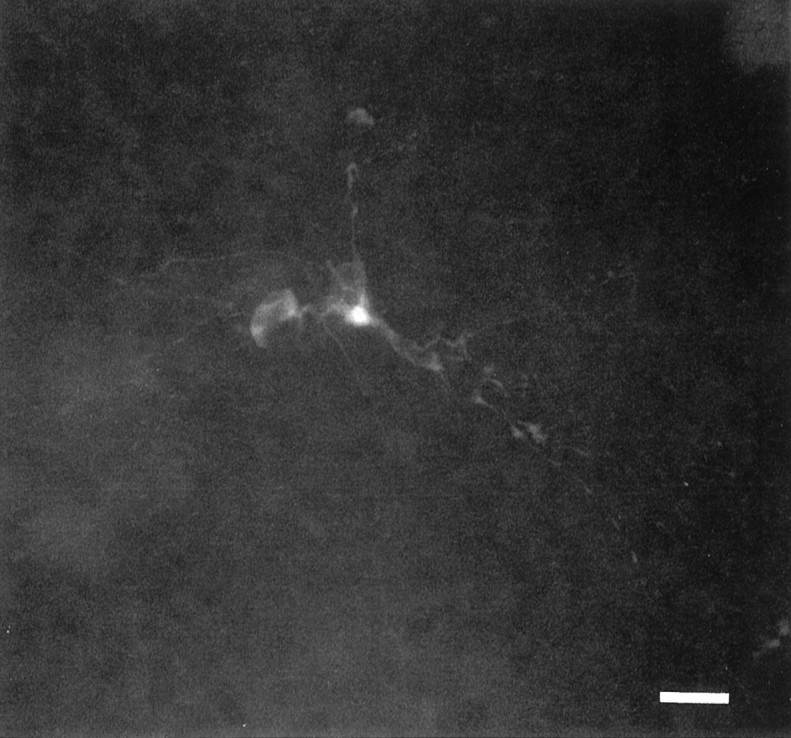
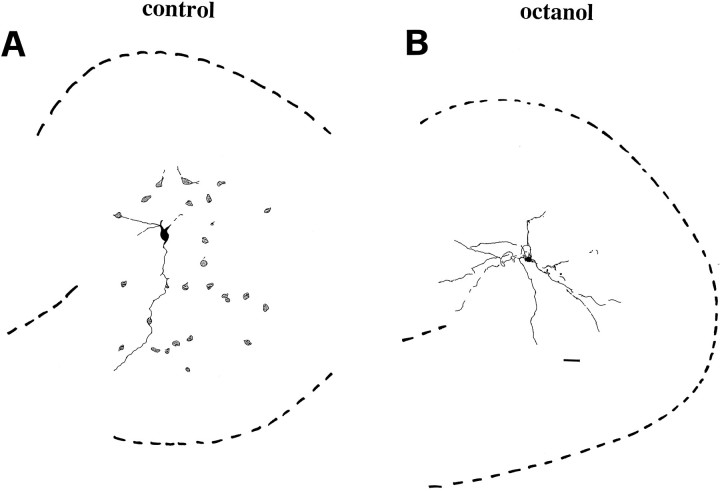
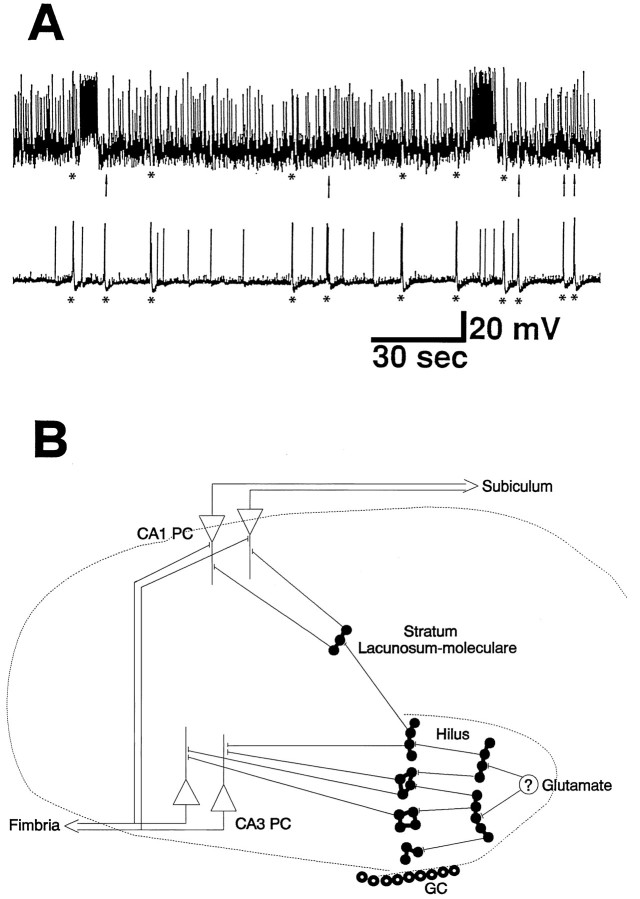
The establishment of synaptic connections and their refinement during development require neural activity. Increasing evidence suggests that spontaneous bursts of neural activity within an immature network are mediated by gamma-aminobutyric acid via a paradoxical excitatory action. Our data show that in the developing hippocampus such synchronous burst activity is generated in the hilar region by transiently coupled cells. These cells have been identified as neuronal elements because they fire action potentials and they are not positive for the glial fibrillary acidic protein staining. Oscillations in hilar cells are "paced" by a hyperpolarization-activated current, with properties of Ih. Coactivated interneurons synchronously release GABA, which via its excitatory action may serve a neurotrophic function during the refinement of hippocampal circuitry.
Figures
References
-
- Altman J, Bayer SA. Migration and distribution of two populations of hippocampal granule cell precursors during the perinatal and postnatal periods. J Comp Neurol. 1990;301:365–381. - PubMed
-
- Amaral DG. A Golgi study of cell types in the hilar region of the hippocampus in the rat. J Comp Neurol. 1978;182:851–914. - PubMed
-
- Andersen P, Bliss VP, Skrede KK. Lamellar organization of hippocampal excitatory pathways. Exp Brain Res. 1971;13:222–238. - PubMed
-
- Barbin G, Pollard H, Gaiarsa JL, Ben Ari Y. Involvement of GABAA receptors in the outgrowth of cultured hippocampal neurons. Neurosci Lett. 1993;152:150–154. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical