

The dynamins: redundant or distinct functions for an expanding family of related GTPases?
- PMID: 9012790
- PMCID: PMC34135
- DOI: 10.1073/pnas.94.2.377
The dynamins: redundant or distinct functions for an expanding family of related GTPases?
Abstract
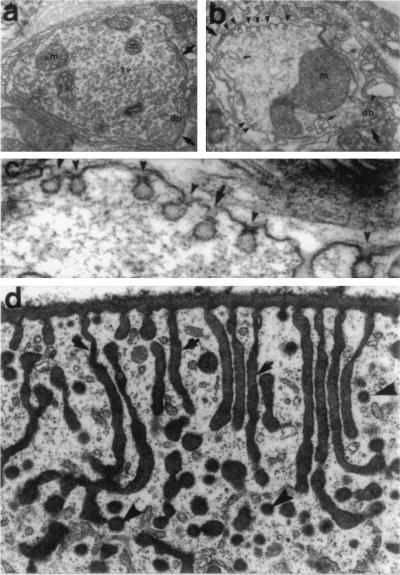
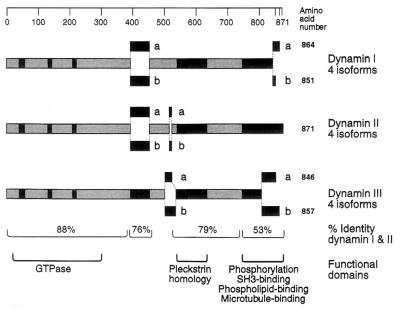
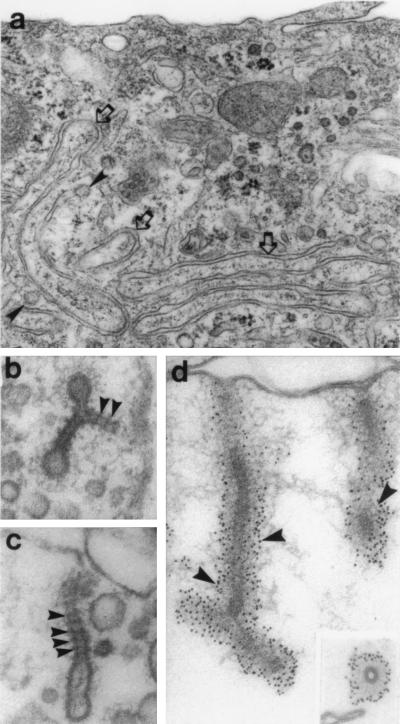
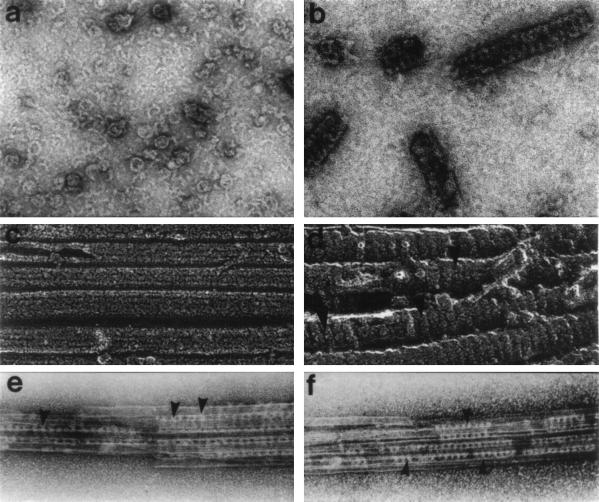
In the 7 years since dynamin was first isolated from bovine brain in search of novel microtubule-based motors, our understanding of this enzyme has expanded significantly. We now know that brain dynamin belongs to a family of large GTPases, which mediate vesicle trafficking. Furthermore, this enzymatic activity is markedly increased through association with microtubules, acidic phospholipids, and certain regulatory proteins that contain Src homology 3 (SH3) domains. From functional, genetic, and cellular manipulations, it is now generally accepted that dynamin participates in the endocytic uptake of receptors, associated ligands, and plasma membrane following an exocytic event. These observations have confirmed at least one function of dynamin that was predicted from seminal studies on a pleiotropic mutant, shibire(ts) (shi(ts)) in Drosophila melanogaster. Of equal interest is the finding that there are multiple dynamin gene products, including two that are expressed in a tissue-specific manner, and they share marked homology with a larger family of distinct but related proteins. Therefore, it is attractive to speculate that the different dynamins may participate in related cellular functions, such as distinct endocytic processes and even secretion. In turn, dynamin could play an important role in cell growth, cell spreading, and neurite outgrowth. The purpose of this review is to enumerate on the expansive dynamin literature and to discuss the nomenclature, expression, and putative functions of this growing and interesting family of proteins.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
