

Probing the environment along the protein import pathways in yeast mitochondria by site-specific photocrosslinking
- PMID: 9012810
- PMCID: PMC19539
- DOI: 10.1073/pnas.94.2.485
Probing the environment along the protein import pathways in yeast mitochondria by site-specific photocrosslinking
Abstract
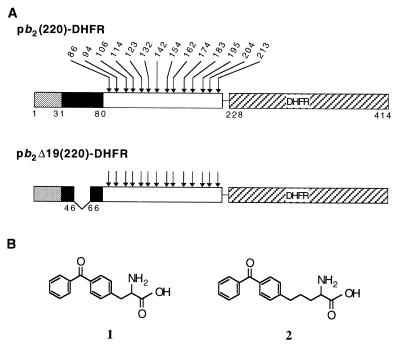
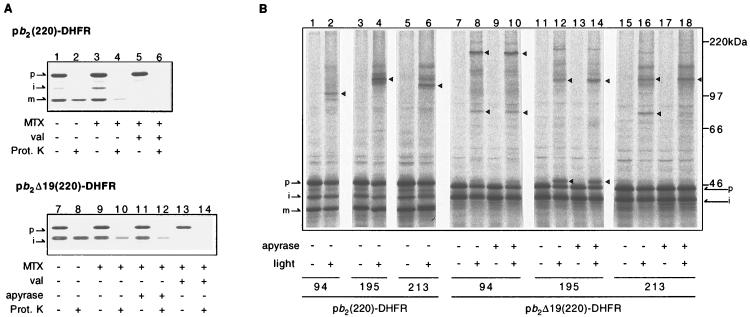
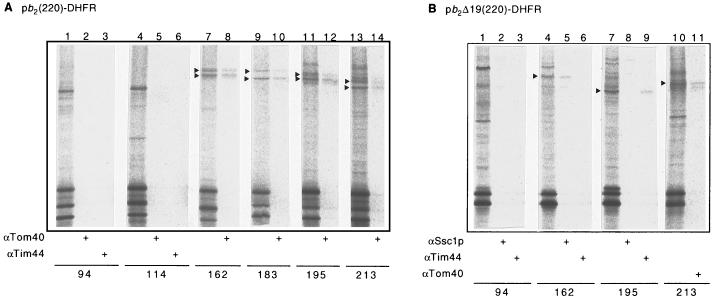
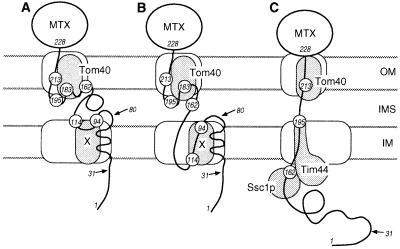
Artificially aminoacylated suppressor tRNAs were used to introduce photoreactive amino acids into model mitochondrial precursor proteins to probe the environment along the protein import pathway. Amino acids with benzophenone side chains of various lengths [DL-2-amino-3-(p-benzoylphenyl)propanoic acid (1) and DL-2-amino-5-(p-benzoylphenyl)pentanoic acid (2)] were incorporated at specific sites throughout the cytochrome b2-dihydrofolate reductase fusion proteins, pb2(220)-DHFR and pb2 delta 19(220)-DHFR, which were destined for the intermembrane space and the matrix in mitochondria, respectively. In vitro import of pb2(220)-DHFR and pb2 delta 19(220)-DHFR bearing 1 or 2 into isolated yeast mitochondria was arrested so that the N terminus reached the intermembrane space or the matrix, respectively, while the DHFR domain remained at the mitochondrial surface. The matrix-targeted pb2 delta 19(220)-DHFR was photocrosslinked to Tom40 in the outer membrane, Tim44 in the inner membrane, and Ssc1p in the matrix, suggesting that the protein has an extended conformation in the import channels. On the other hand, incorporation of 2 at various positions in the 50-residue segment of intermembrane-space-targeted pb2(220)-DHFR gave photocrosslinks only to Tom40, suggesting that the segment is not in an extended conformation, but localized near Tom40. The N-terminal portion of pb2(220)-DHFR, but not pb2 delta 19(220)-DHFR, was photocrosslinked to an as-yet-unidentified mitochondrial component to generate a 95-kDa crosslinked product.
Figures
References
-
- Lill R, Nargang F E, Neupert W. Curr Opin Cell Biol. 1996;8:505–512. - PubMed
-
- Schatz G, Dobberstein B. Science. 1996;271:1519–1526. - PubMed
-
- Mihara K, Omura T. Trends Cell Biol. 1996;6:104–108. - PubMed
-
- Lill R, Neupert W. Trends Cell Biol. 1996;6:56–61. - PubMed
-
- Pfanner N, Craig E A, Meijer M. Trends Biochem Sci. 1994;19:368–372. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
