

Interferon-alpha/beta inhibition of interleukin 12 and interferon-gamma production in vitro and endogenously during viral infection
- PMID: 9012836
- PMCID: PMC19565
- DOI: 10.1073/pnas.94.2.634
Interferon-alpha/beta inhibition of interleukin 12 and interferon-gamma production in vitro and endogenously during viral infection
Abstract
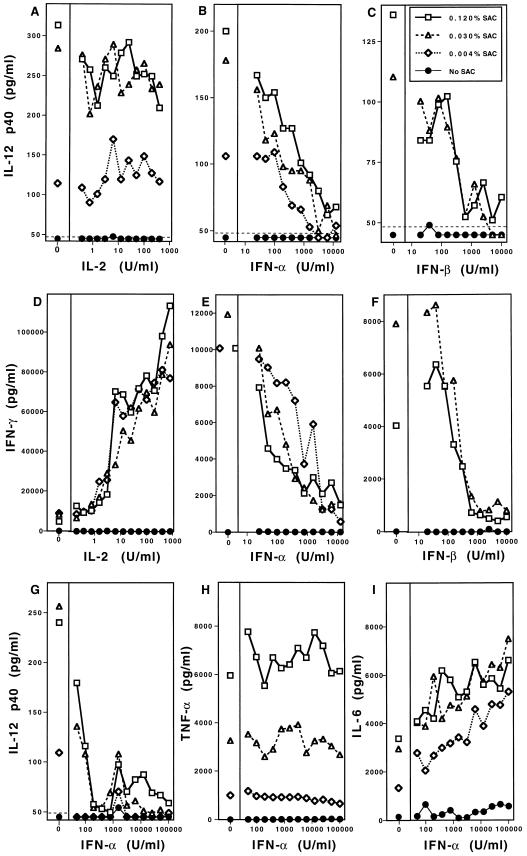
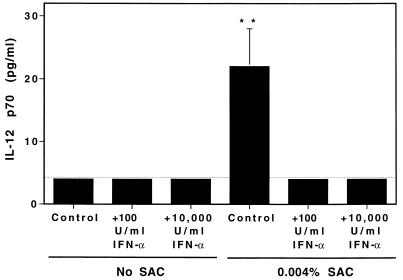
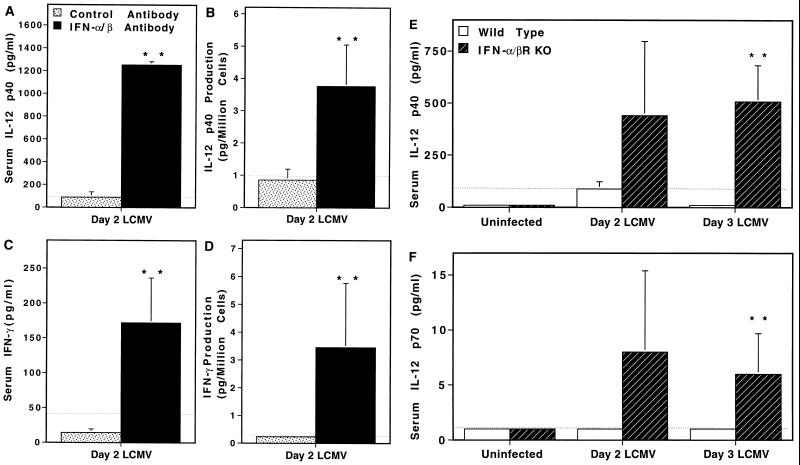
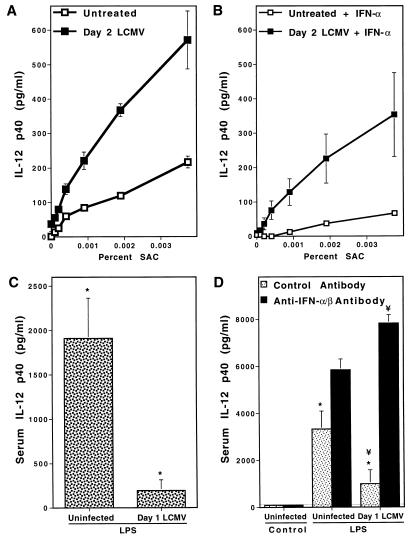
Interferon (IFN)-alpha/beta-mediated negative regulation of interleukin 12 (IL-12) and IFN-gamma proteins is reported here. Both IFN-alpha and IFN-beta inhibited fixed Staphylococcus aureus Cowan strain induction of IL-12 and IFN-gamma production by mouse splenic leukocytes in culture. Extended studies with IFN-alpha demonstrated that inhibition was at the level of biologically active IL-12 p70. Effects were selective, as induction of tumor necrosis factor was unaffected and induction of IL-6 was enhanced. Neutralization of IFN-alpha/beta expressed endogenously during infections with murine cytomegalovirus (MCMV) enhanced early IL-12 and IFN-gamma protein production. Furthermore, during infections of mice with lymphocytic choriomeningitis virus (LCMV), this treatment revealed a previously undetected early IL-12 and IFN-gamma protein expression, and mice deficient in IFN-alpha/beta receptor function, but not control mice, also expressed endogenous LCMV-induced IL-12. The effects of IFN-alpha/beta neutralization on production of IL-12 and IFN-gamma during the viral infections were detected in both serum samples and medium conditioned with splenic leukocytes isolated from infected animals. In vitro studies demonstrated that splenic leukocytes isolated from LCMV-infected mice were primed to produce IL-12 in response to stimulation with Staphylococcus aureus Cowan strain, but that this responsiveness was sensitive to added IFN-alpha. Moreover, endogenous IFN-alpha/beta induced by LCMV inhibited in vivo lipopolysaccharide stimulation of IL-12 production. These results demonstrate a new pathway for regulating cytokine responses, and suggest a mechanism for inhibition of IL-12-dependent immune responses during viral infections.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
