

Demonstration of mechanical connections between integrins, cytoskeletal filaments, and nucleoplasm that stabilize nuclear structure
- PMID: 9023345
- PMCID: PMC19602
- DOI: 10.1073/pnas.94.3.849
Demonstration of mechanical connections between integrins, cytoskeletal filaments, and nucleoplasm that stabilize nuclear structure
Abstract
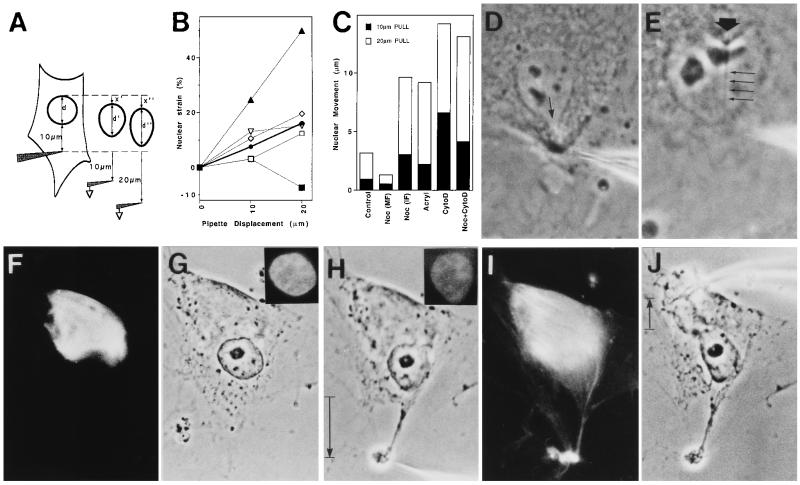
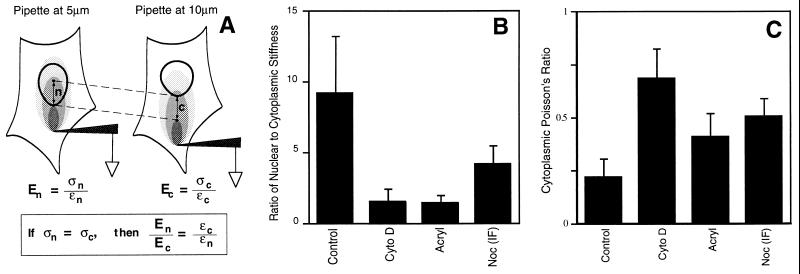
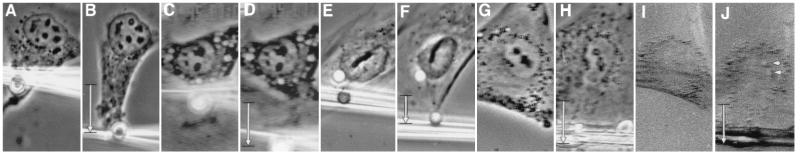
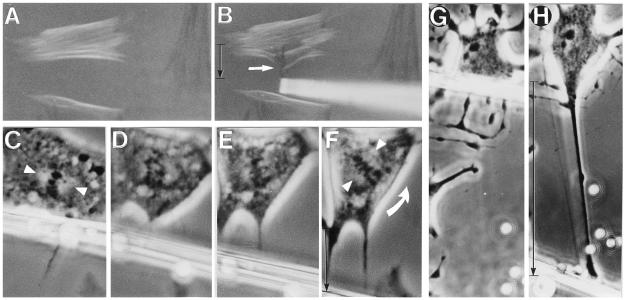
We report here that living cells and nuclei are hard-wired such that a mechanical tug on cell surface receptors can immediately change the organization of molecular assemblies in the cytoplasm and nucleus. When integrins were pulled by micromanipulating bound microbeads or micropipettes, cytoskeletal filaments reoriented, nuclei distorted, and nucleoli redistributed along the axis of the applied tension field. These effects were specific for integrins, independent of cortical membrane distortion, and were mediated by direct linkages between the cytoskeleton and nucleus. Actin microfilaments mediated force transfer to the nucleus at low strain; however, tearing of the actin gel resulted with greater distortion. In contrast, intermediate filaments effectively mediated force transfer to the nucleus under both conditions. These filament systems also acted as molecular guy wires to mechanically stiffen the nucleus and anchor it in place, whereas microtubules acted to hold open the intermediate filament lattice and to stabilize the nucleus against lateral compression. Molecular connections between integrins, cytoskeletal filaments, and nuclear scaffolds may therefore provide a discrete path for mechanical signal transfer through cells as well as a mechanism for producing integrated changes in cell and nuclear structure in response to changes in extracellular matrix adhesivity or mechanics.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
