

Oncogenic H-ras stimulates tumor angiogenesis by two distinct pathways
- PMID: 9023347
- PMCID: PMC19604
- DOI: 10.1073/pnas.94.3.861
Oncogenic H-ras stimulates tumor angiogenesis by two distinct pathways
Abstract
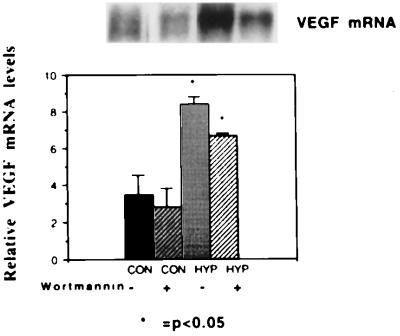
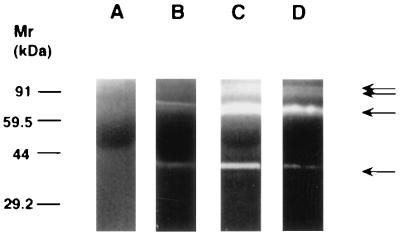
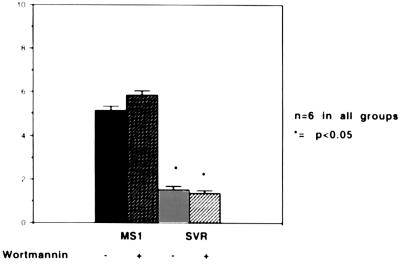
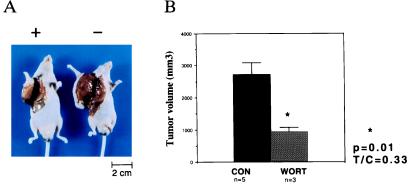
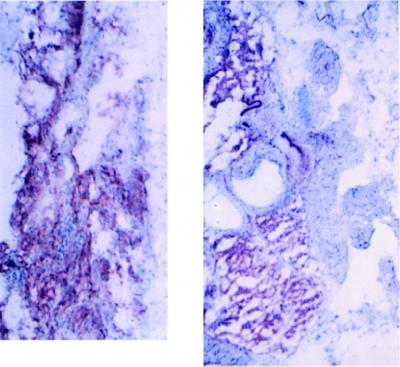
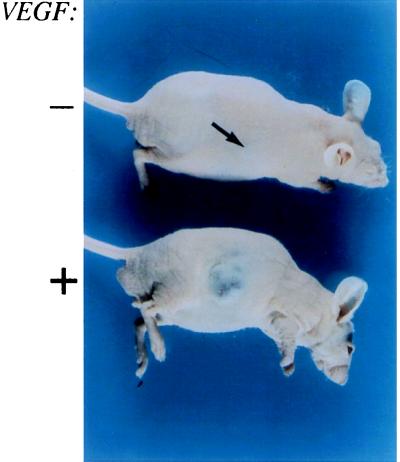
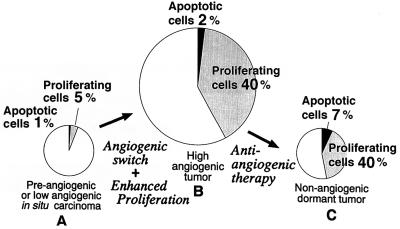
The switch from a quiescent tumor to an invasive tumor is accompanied by the acquisition of angiogenic properties. This phenotypic change likely requires a change in the balance of angiogenic stimulators and angiogenic inhibitors. The nature of the angiogenic switch is not known. Here, we show that introduction of activated H-ras into immortalized endothelial cells is capable of activating the angiogenic switch. Angiogenic switching is accompanied by up-regulation of vascular endothelial growth factor and matrix metalloproteinase (MMP) bioactivity and downregulation of tissue inhibitor of MMP. Furthermore, we show that inhibition of phosphatidylinositol-3-kinase leads to partial inhibition of tumor angiogenesis, thus demonstrating that activated H-ras activates tumor angiogenesis through two distinct pathways. Finally, we show evidence for two forms of tumor dormancy.
Figures
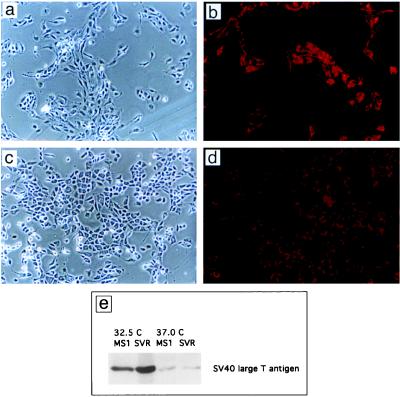
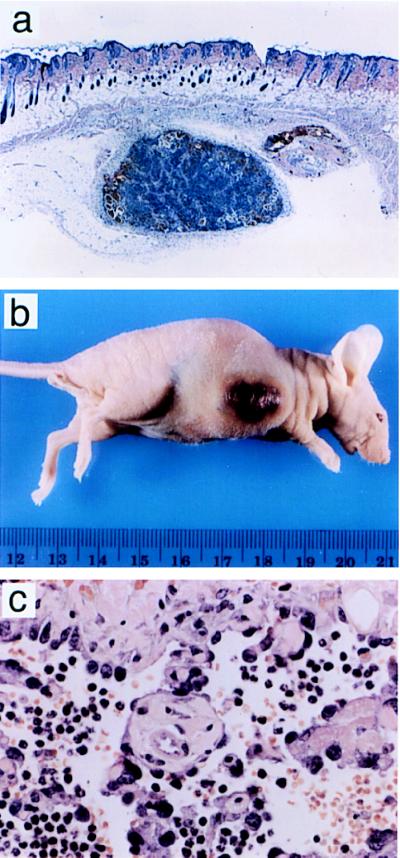
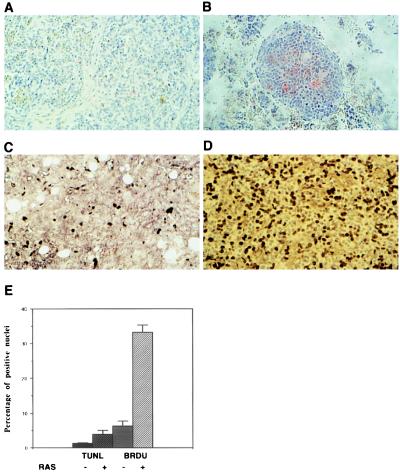
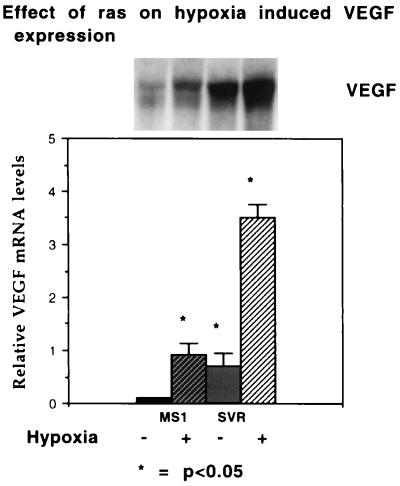
References
-
- Folkman J, Hanahan D. Cell. 1996;86:353–364. - PubMed
-
- Folkman J, Watson K, Ingber D, Hanahan D. Nature (London) 1989;339:58–61. - PubMed
-
- Kandel J, Bossy-Wetzel E, Radvanyi F, Klagsbrun M, Folkman J, Hanahan D. Cell. 1991;66:1095–1104. - PubMed
-
- Moses M A, Sudhalter J, Langer R. Science. 1990;248:1408–1410. - PubMed
-
- Liotta L A, Steeg P S, Stetler-Stevenson W G. Cell. 1991;64:327–336. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
