

Functional nonequality of the cardiac and skeletal ryanodine receptors
- PMID: 9023375
- PMCID: PMC19632
- DOI: 10.1073/pnas.94.3.1019
Functional nonequality of the cardiac and skeletal ryanodine receptors
Abstract
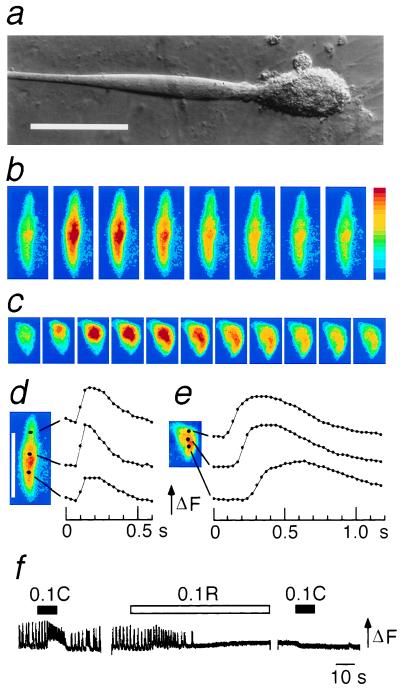
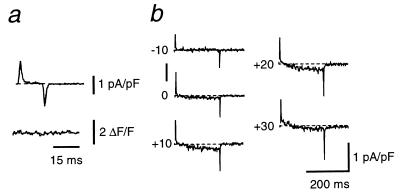
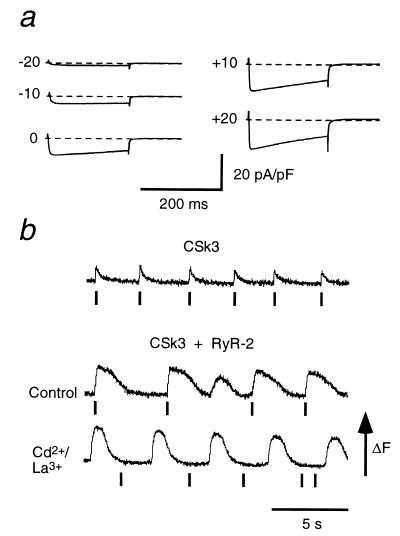
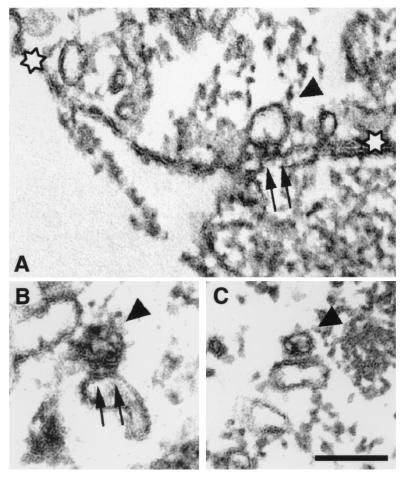
Dihydropyridine receptors (DHPRs), which are voltage-gated Ca2+ channels, and ryanodine receptors (RyRs), which are intracellular Ca2+ release channels, are expressed in diverse cell types, including skeletal and cardiac muscle. In skeletal muscle, there appears to be reciprocal signaling between the skeletal isoforms of both the DHPR and the RyR (RyR-1), such that Ca2+ release activity of RyR-1 is controlled by the DHPR and Ca2+ channel activity of the DHPR is controlled by RyR-1. Dyspedic skeletal muscle cells, which do not express RyR-1, lack excitation-contraction coupling and have an approximately 30-fold reduction in L-type Ca2+ current density. Here we have examined the ability of the predominant cardiac and brain RyR isoform, RyR-2, to substitute for RyR-1 in interacting with the skeletal DHPR. When RyR-2 is expressed in dyspedic muscle cells, it gives rise to spontaneous intracellular Ca2+ oscillations and supports Ca2+ entry-induced Ca2+ release. However, unlike RyR-1, the expressed RyR-2 does not increase the Ca2+ channel activity of the DHPR, nor is the gating of RyR-2 controlled by the skeletal DHPR. Thus, the ability to participate in skeletal-type reciprocal signaling appears to be a unique feature of RyR-1.
Figures
References
-
- Hofmann F, Biel M, Flockerzi V. Annu Rev Neurosci. 1994;17:399–418. - PubMed
-
- Tanabe T, Takeshima H, Mikami A, Flockerzi V, Takahashi H, Kangawa K, Kojima M, Matsuo H, Hirose T, Numa S. Nature (London) 1987;328:313–318. - PubMed
-
- Mikami A, Imoto K, Tanabe T, Niidome T, Mori Y, Takeshima H, Narumiya S, Numa S. Nature (London) 1989;340:230–233. - PubMed
-
- Williams M E, Feldman D H, McCue A F, Brenner R, Velicelebi G, Ellis S B, Harpold M M. Neuron. 1992;8:71–84. - PubMed
-
- McPherson P S, Campbell K P. J Biol Chem. 1993;268:13765–13768. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
