

Interphase nuclei of many mammalian cell types contain deep, dynamic, tubular membrane-bound invaginations of the nuclear envelope
- PMID: 9024685
- PMCID: PMC2134289
- DOI: 10.1083/jcb.136.3.531
Interphase nuclei of many mammalian cell types contain deep, dynamic, tubular membrane-bound invaginations of the nuclear envelope
Abstract
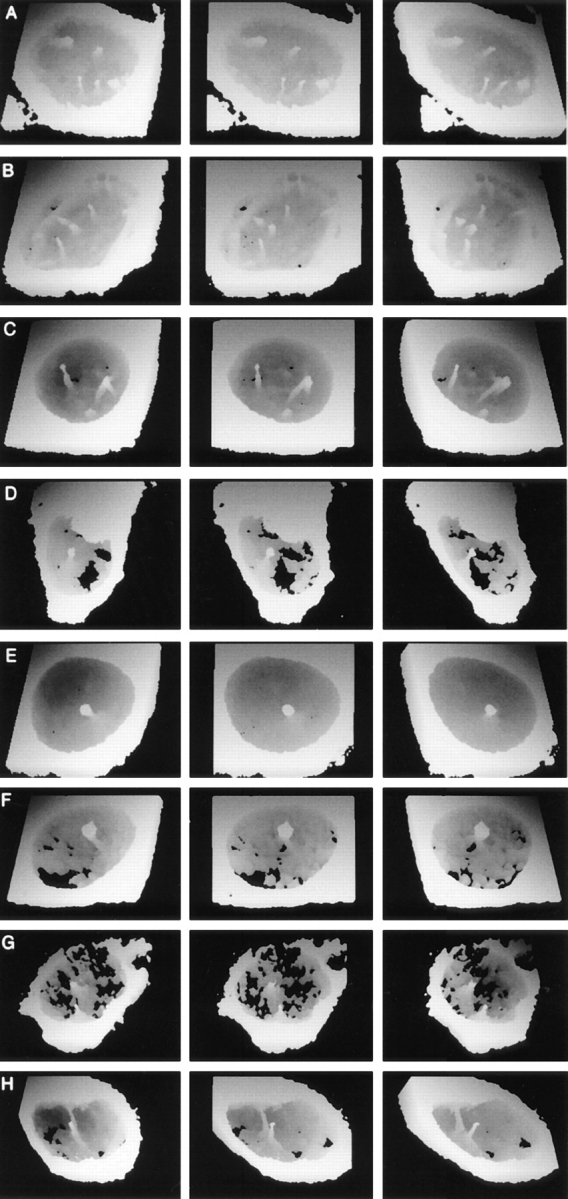
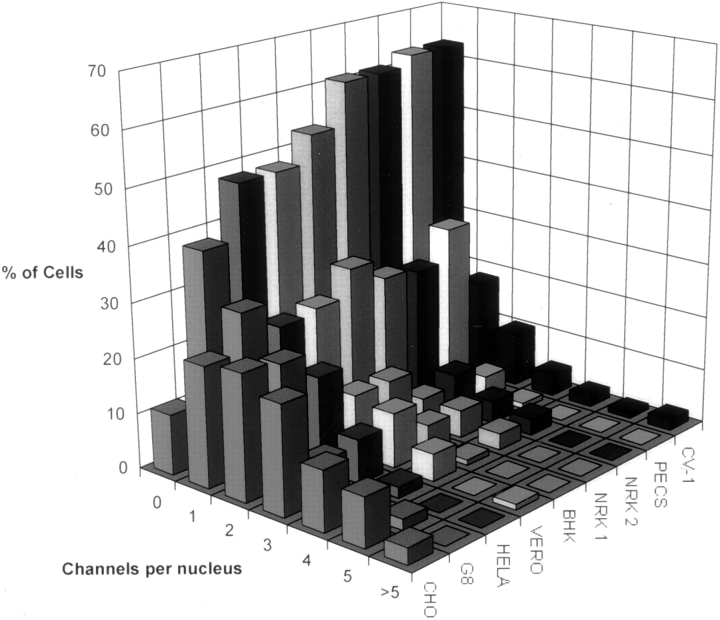
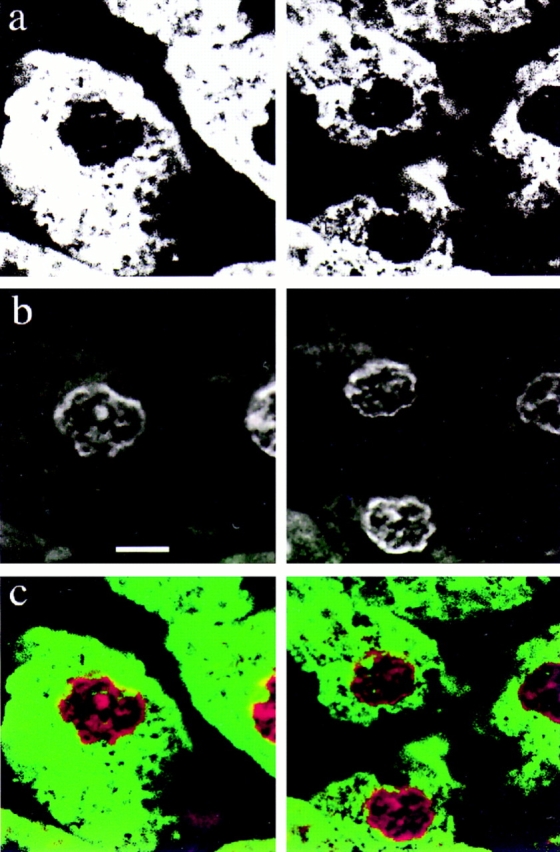
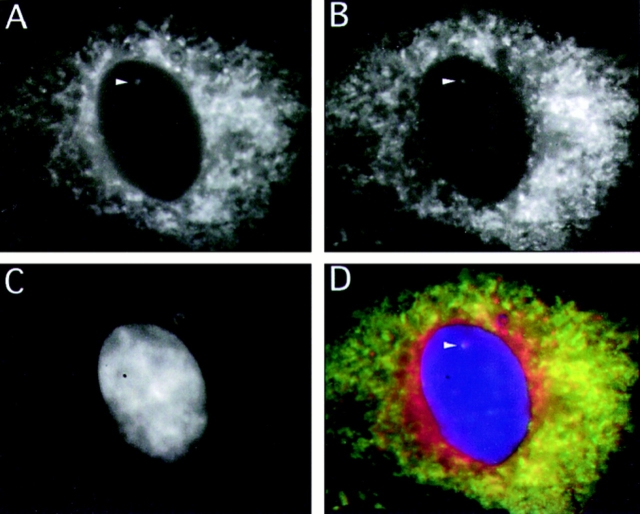
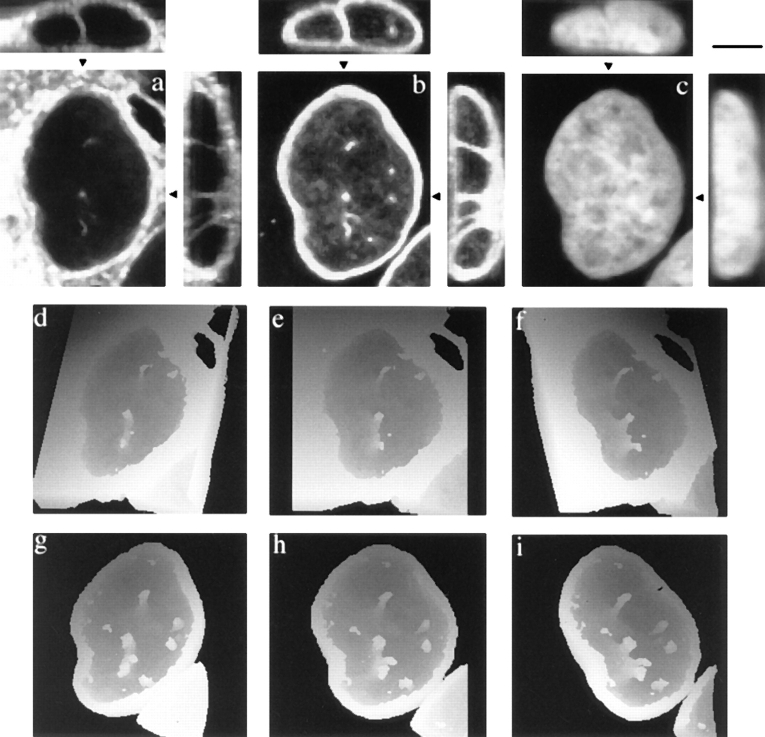
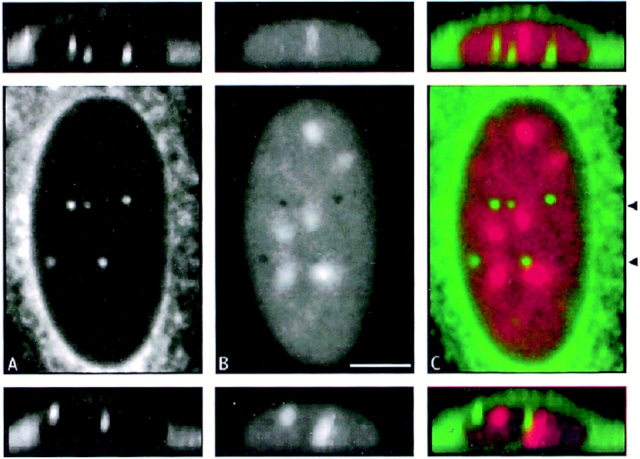
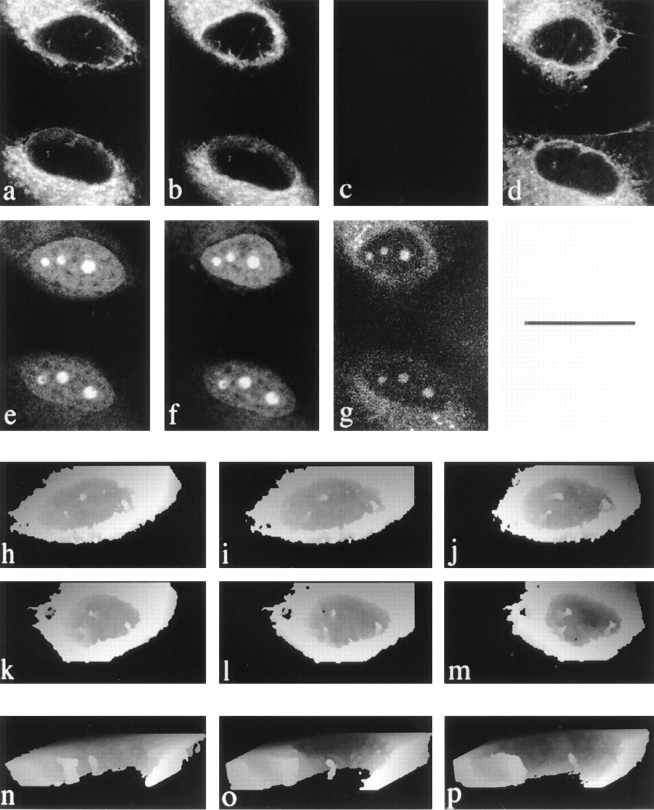
The nuclear envelope consists of a double-membraned extension of the rough endoplasmic reticulum. In this report we describe long, dynamic tubular channels, derived from the nuclear envelope, that extend deep into the nucleoplasm. These channels show cell-type specific morphologies ranging from single short stubs to multiple, complex, branched structures. Some channels transect the nucleus entirely, opening at two separate points on the nuclear surface, while others terminate at or close to nucleoli. These channels are distinct from other topological features of the nuclear envelope, such as lobes or folds. The channel wall consists of two membranes continuous with the nuclear envelope, studded with features indistinguishable from nuclear pore complexes, and decorated on the nucleoplasmic surface with lamins. The enclosed core is continuous with the cytoplasm, and the lumenal space between the membranes contains soluble ER-resident proteins (protein disulphide isomerase and glucose-6-phosphatase). Nuclear channels are also found in live cells labeled with the lipophilic dye DiOC6. Time-lapse imaging of DiOC6-labeled cells shows that the channels undergo changes in morphology and spatial distribution within the interphase nucleus on a timescale of minutes. The presence of a cytoplasmic core and nuclear pore complexes in the channel walls suggests a possible role for these structures in nucleo-cytoplasmic transport. The clear association of a subset of these structures with nucleoli would also be consistent with such a transport role.
Figures
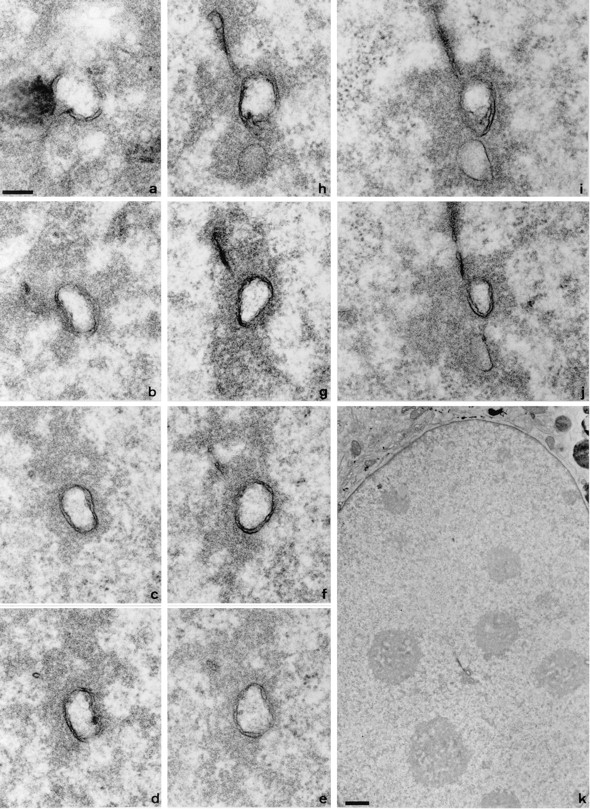
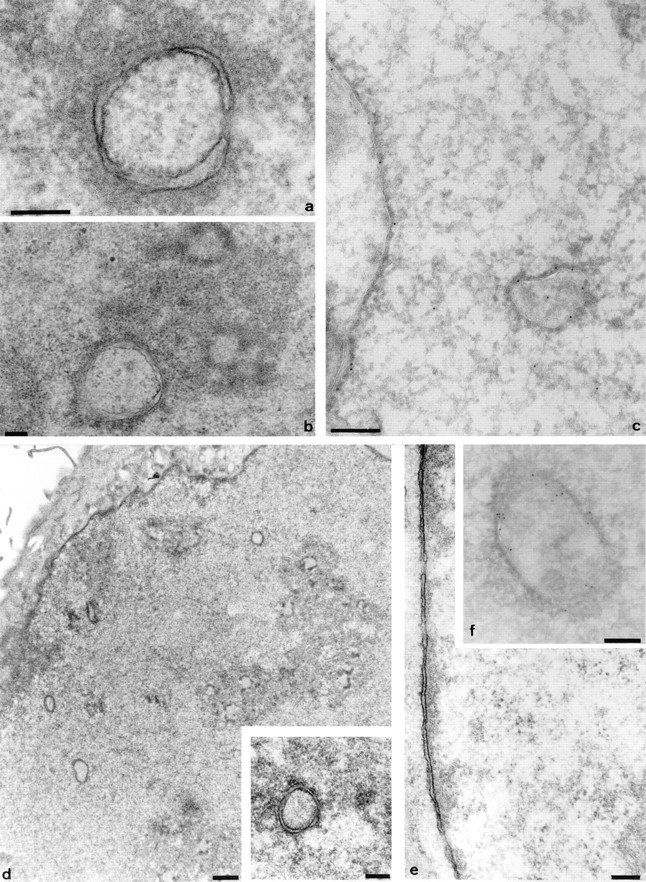
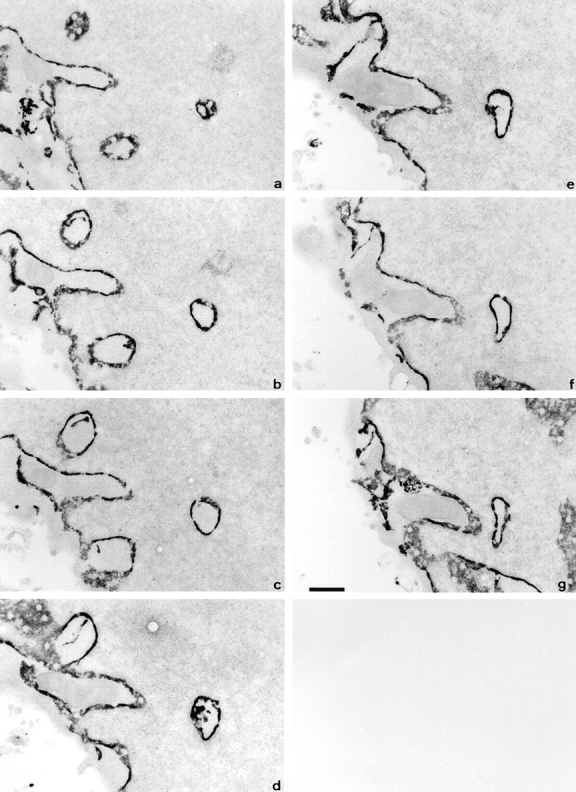
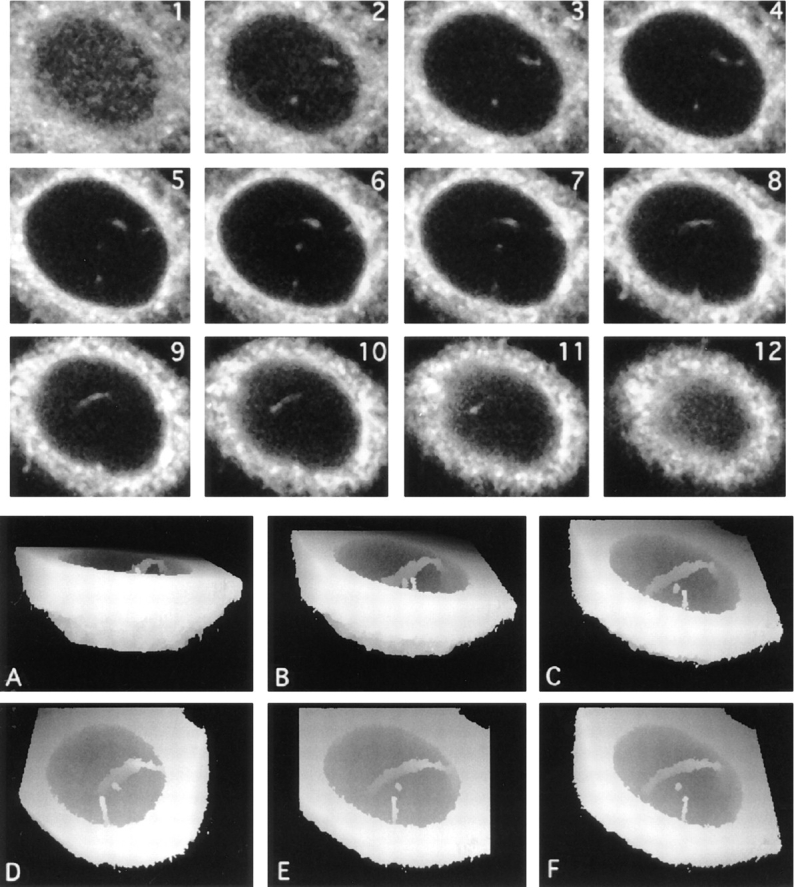
References
-
- Banfalvi G, Wiegant J, Sarkar N, van-Duijn P. Immunofluorescent visualization of DNA replication sites within nuclei of Chinese hamster ovary cells. Histochemistry. 1989;93:81–86. - PubMed
-
- Bourgeois CA, Hemon D, Bouteille M. Structural relationship between the nucleolus and the nuclear envelope. J Ultrastruct Res. 1979;68:328–340. - PubMed
-
- Bowers CW, Dahm LM. Buffers for cell culture. Trends Cell Biol. 1993;3:76–77. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
