

A cytosolic serine endopeptidase from Trypanosoma cruzi is required for the generation of Ca2+ signaling in mammalian cells
- PMID: 9024691
- PMCID: PMC2134300
- DOI: 10.1083/jcb.136.3.609
A cytosolic serine endopeptidase from Trypanosoma cruzi is required for the generation of Ca2+ signaling in mammalian cells
Abstract
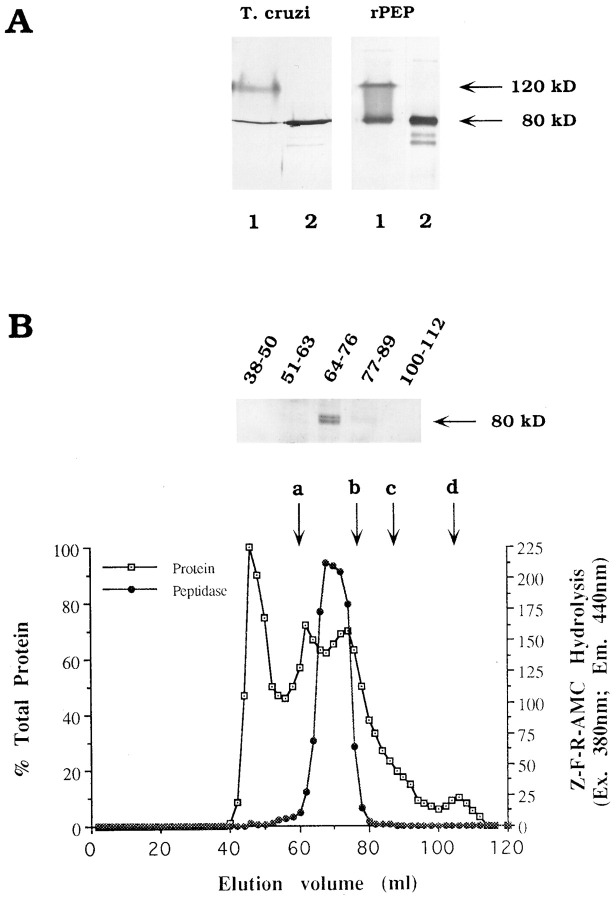
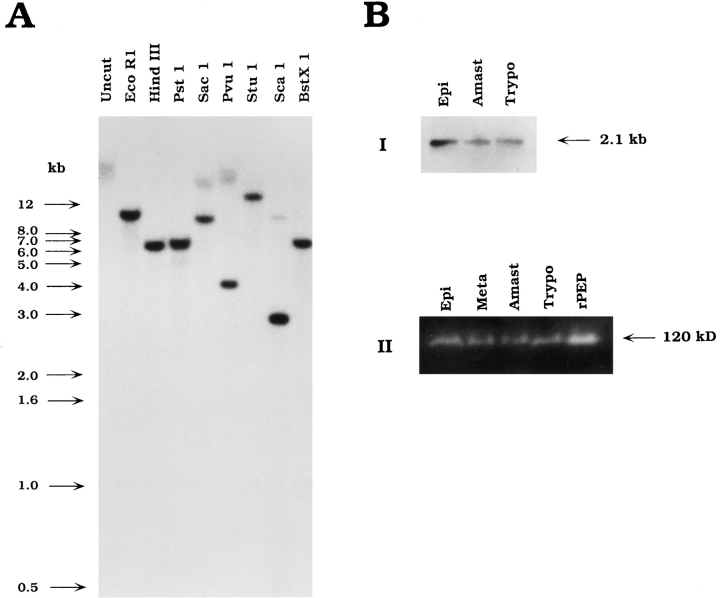
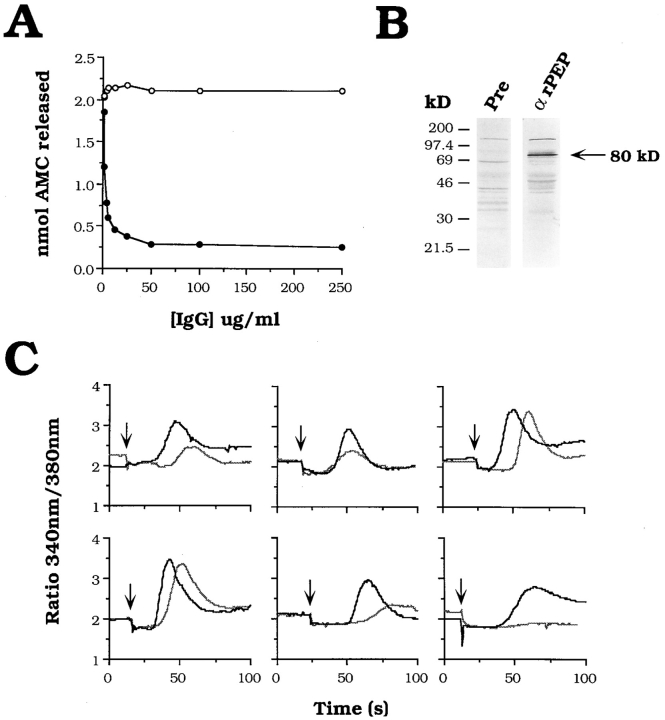
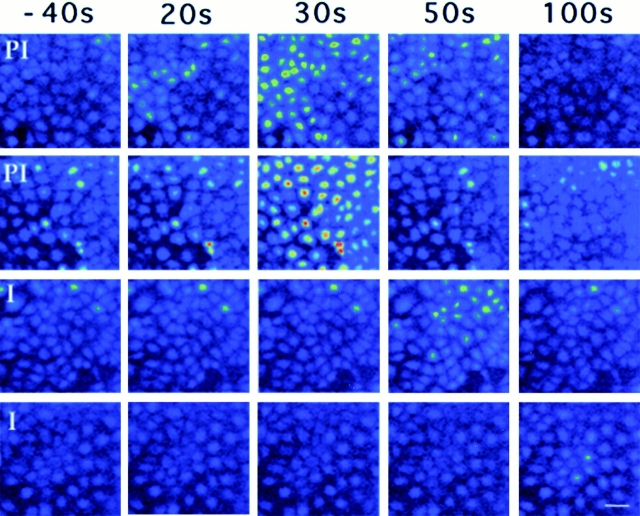
An early event in the Trypanosoma cruzi cell invasion process, the recruitment of host lysosomes, led us to investigate the involvement of signal transduction. Infective trypomastigotes were found to contain a soluble Ca2+-signaling activity for mammalian cells that is sensitive to protease inhibitors. Inhibitor and substrate utilization profiles were used to purify a candidate peptidase for involvement in this process, from which we isolated a full-length cDNA clone. The sequence revealed a novel enzyme, denominated T. cruzi oligopeptidase B, which is homologous to members of the prolyl oligopeptidase family of serine hydrolases, known to participate in the maturation of biologically active peptides. The T. cruzi oligopeptidase B was expressed as a fully active product in Escherichia coli, and antibodies to the recombinant enzyme inhibited both peptidase activity and Ca2+ signaling induced in normal rat kidney cells by trypomastigote extracts. Our data suggest that the T. cruzi oligopeptidase B participates in processing events in the cytoplasm of the parasites, generating a factor with Ca2+-signaling activity for mammalian cells.
Figures

References
-
- Andrews NW, Hong KS, Robbins ES, Nussenzweig V. Stagespecific surface antigens expressed during the morphogenesis of vertebrate forms of Trypanosoma cruzi. . Exp Parasitol. 1987;64:474–484. - PubMed
-
- Ashall F. Characterization of an alkaline peptidase of Trypanosoma cruziand other trypanosomatids. Mol Biochem Parasitol. 1990;38:77–87. - PubMed
-
- Barrett AJ, Rawlings ND. Oligopeptidases, and the emergence of the prolyl oligopeptidase family. Biol Chem Hoppe-Seyler. 1992;373:353–360. - PubMed
-
- Bliska JB, Galan JE, Falkow S. Signal transduction in the mammalian cell during bacterial attachment and entry. Cell. 1993;73:903–920. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
