

Threshold levels of fluid shear promote leukocyte adhesion through selectins (CD62L,P,E)
- PMID: 9024700
- PMCID: PMC2134292
- DOI: 10.1083/jcb.136.3.717
Threshold levels of fluid shear promote leukocyte adhesion through selectins (CD62L,P,E)
Erratum in
- J Cell Biol 1997 Apr 7;137(1):261
Abstract
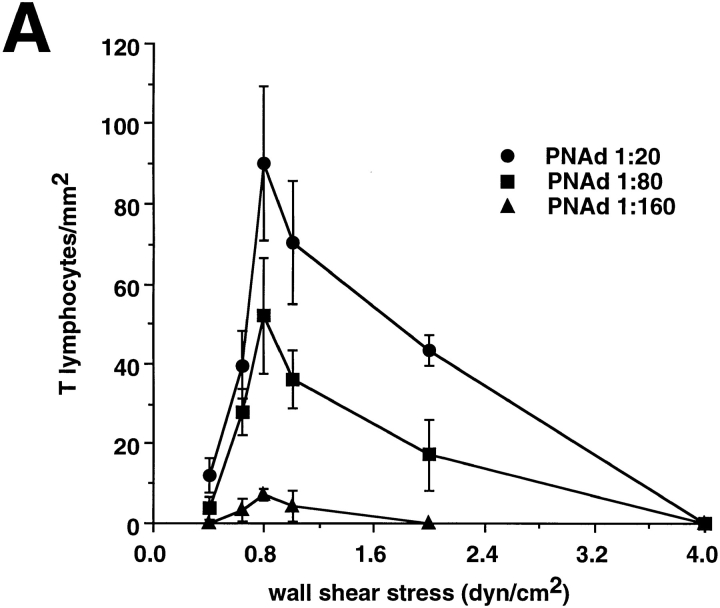
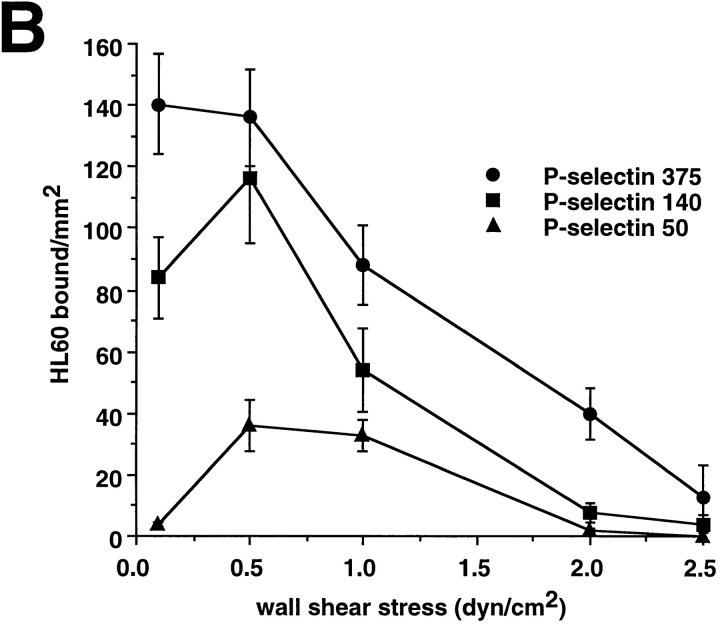
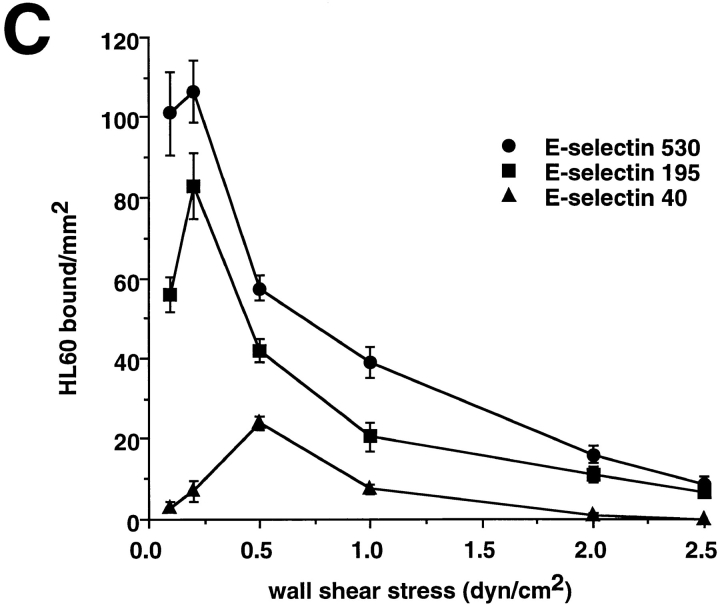
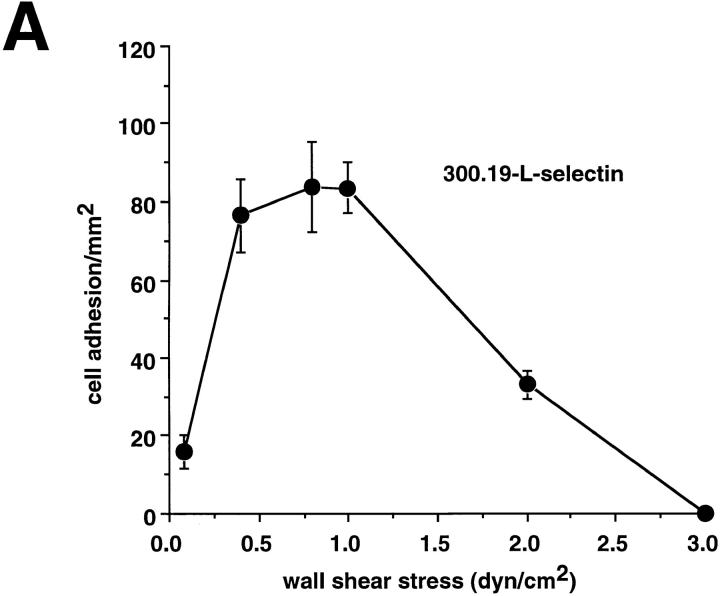
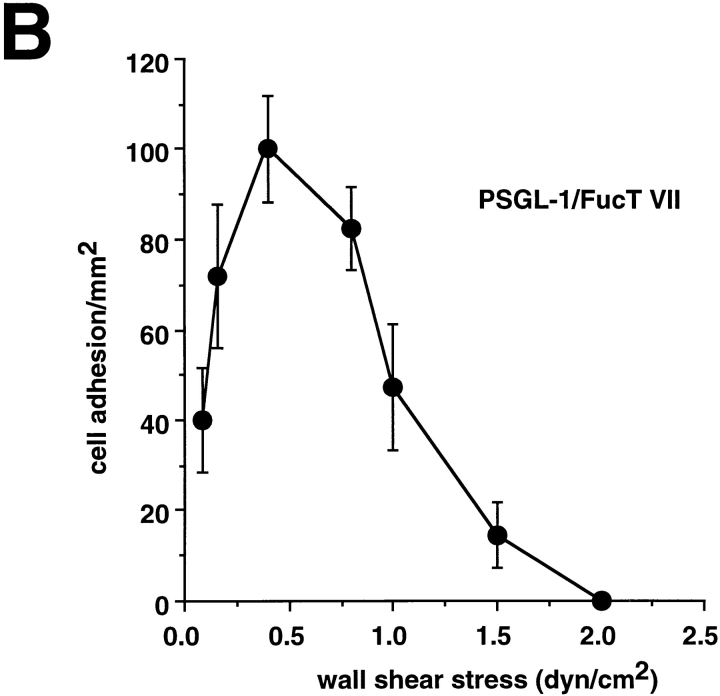
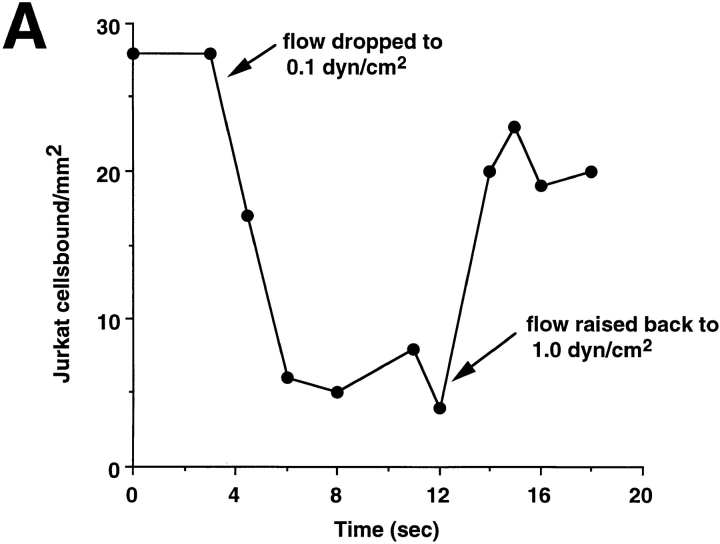
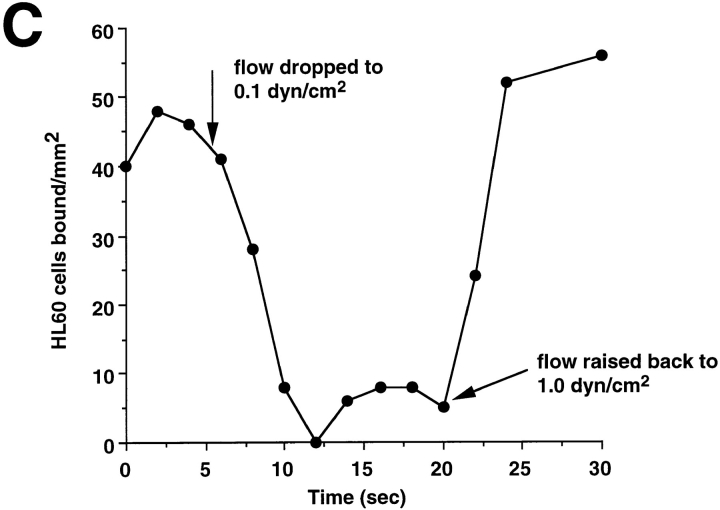
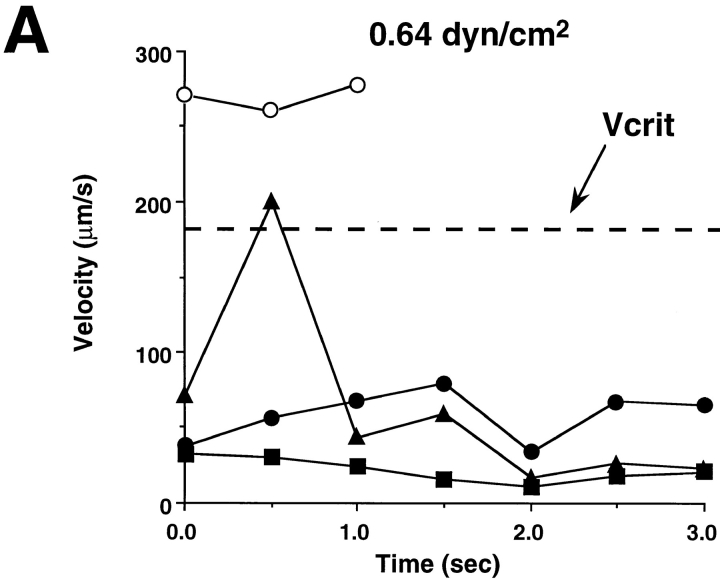
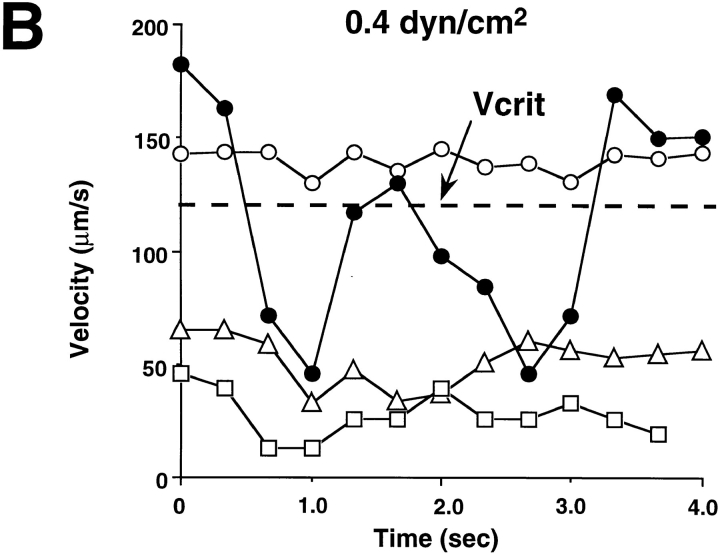
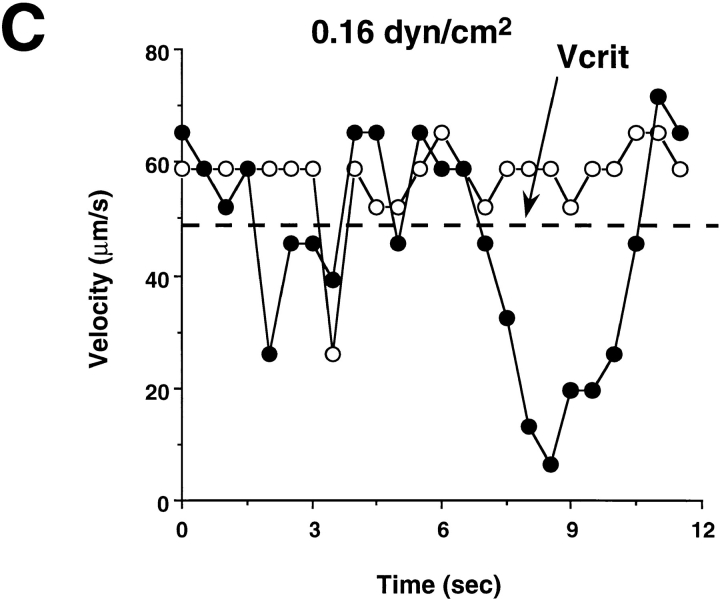
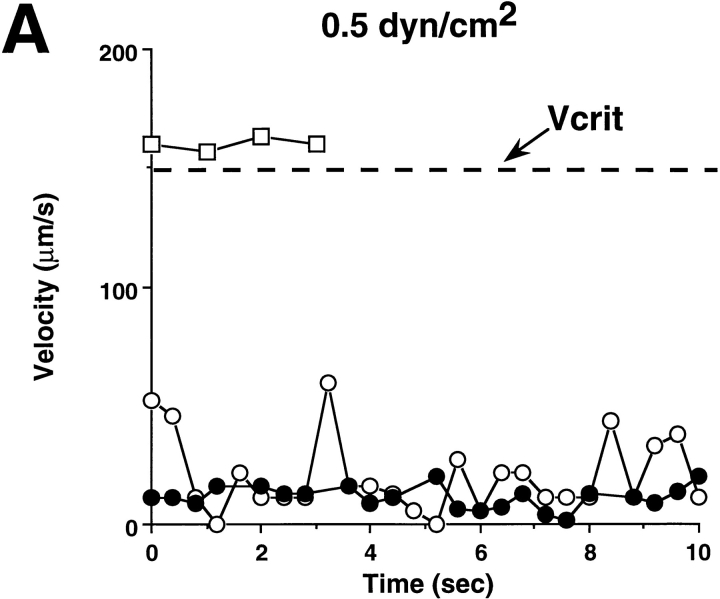
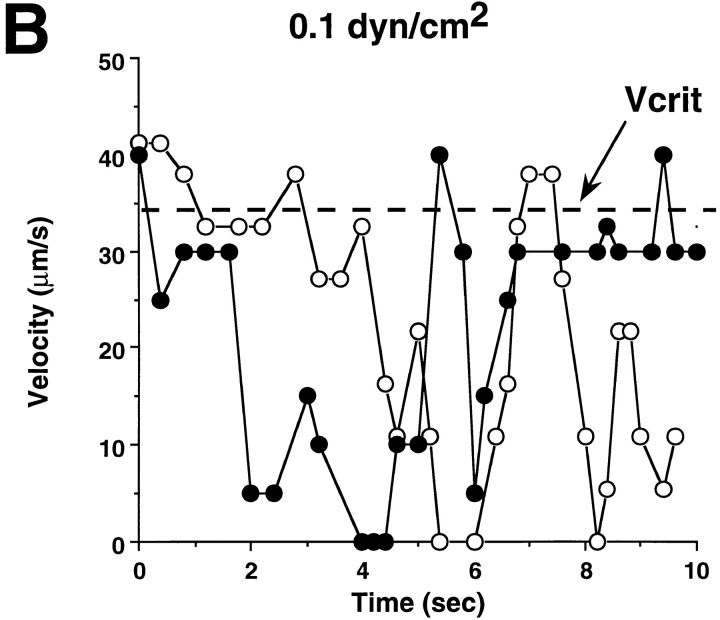
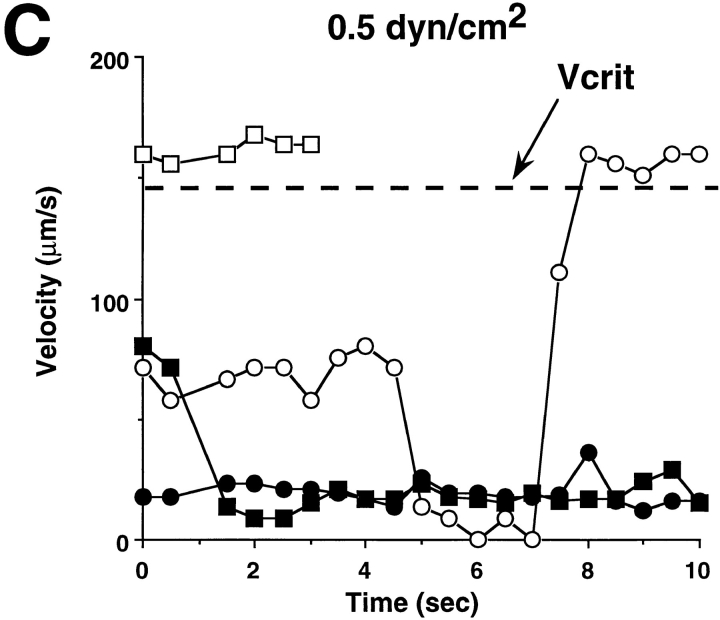
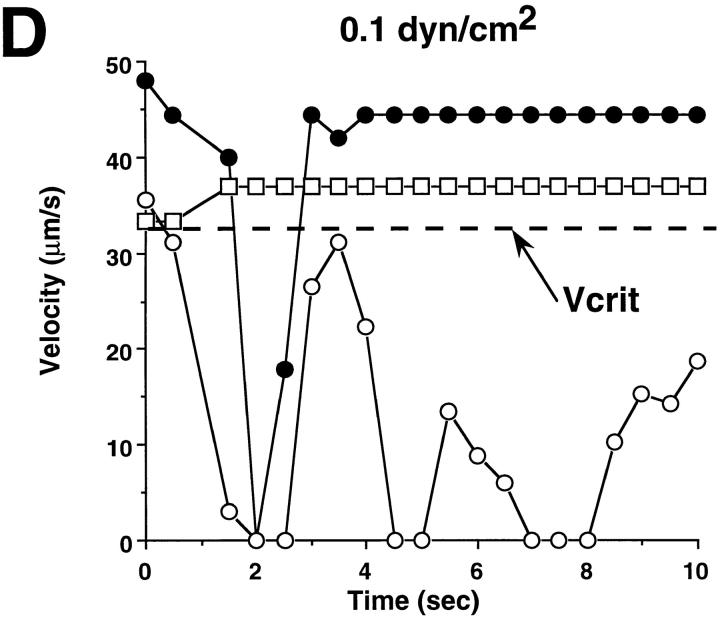
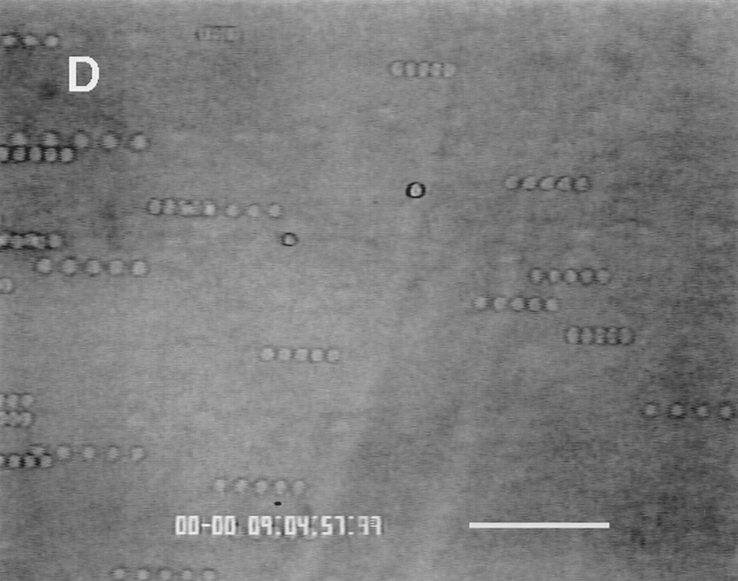
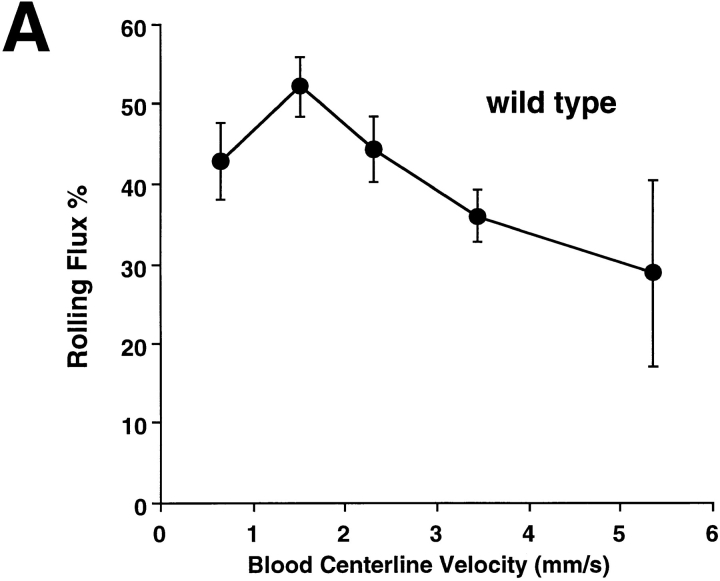
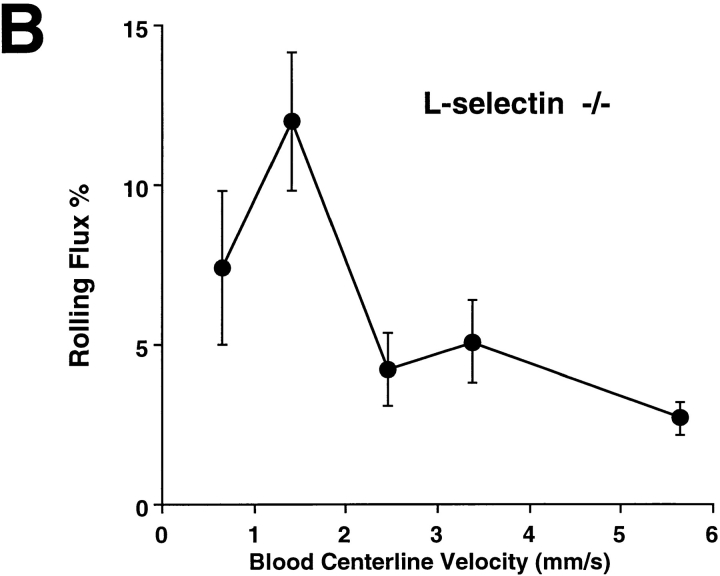
Leukocyte adhesion through L-selectin to peripheral node addressin (PNAd, also known as MECA-79 antigen), an L-selectin ligand expressed on high endothelial venules, has been shown to require a minimum level of fluid shear stress to sustain rolling interactions (Finger, E.B., K.D. Puri, R. Alon, M.B. Lawrence, V.H. von Andrian, and T.A. Springer. 1996. Nature (Lond.). 379:266-269). Here, we show that fluid shear above a threshold of 0.5 dyn/cm2 wall shear stress significantly enhances HL-60 myelocyte rolling on P- and E-selectin at site densities of 200/microm2 and below. In addition, gravitational force is sufficient to detach HL-60 cells from P- and E-selectin substrates in the absence, but not in the presence, of flow. It appears that fluid shear-induced torque is critical for the maintenance of leukocyte rolling. K562 cells transfected with P-selectin glycoprotein ligand-1, a ligand for P-selectin, showed a similar reduction in rolling on P-selectin as the wall shear stress was lowered below 0.5 dyn/cm2. Similarly, 300.19 cells transfected with L-selectin failed to roll on PNAd below this level of wall shear stress, indicating that the requirement for minimum levels of shear force is not cell type specific. Rolling of leukocytes mediated by the selectins could be reinitiated within seconds by increasing the level of wall shear stress, suggesting that fluid shear did not modulate receptor avidity. Intravital microscopy of cremaster muscle venules indicated that the leukocyte rolling flux fraction was reduced at blood centerline velocities less than 1 mm/s in a model in which rolling is mediated by L- and P-selectin. Similar observations were made in L-selectin-deficient mice in which leukocyte rolling is entirely P-selectin dependent. Leukocyte adhesion through all three selectins appears to be significantly enhanced by a threshold level of fluid shear stress.
Figures


References
-
- Alon R, Hammer DA, Springer TA. Lifetime of the P-selectincarbohydrate bond and its response to tensile force in hydrodynamic flow. Nature (Lond) 1995a;374:539–542. - PubMed
-
- Arbones ML, Ord DC, Ley K, Ratech H, Curry-Maynard C, Otten G, Capon DJ, Tedder TF. Lymphocyte homing and leukocyte rolling and migration are impaired in L-selectin-deficient mice. Immunity. 1994;1:247–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
