

Opiate receptor knockout mice define mu receptor roles in endogenous nociceptive responses and morphine-induced analgesia
- PMID: 9037090
- PMCID: PMC19828
- DOI: 10.1073/pnas.94.4.1544
Opiate receptor knockout mice define mu receptor roles in endogenous nociceptive responses and morphine-induced analgesia
Abstract
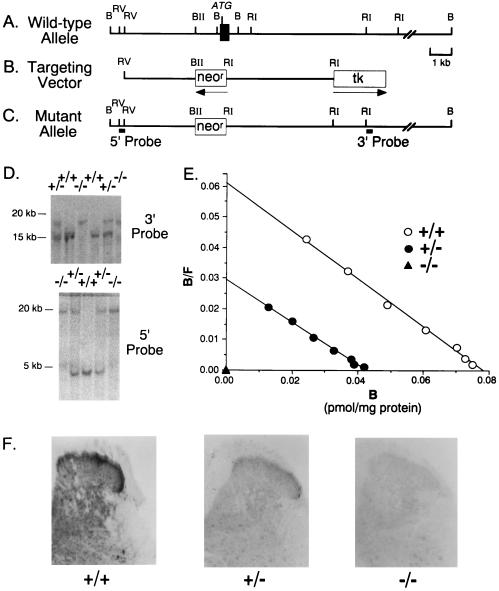
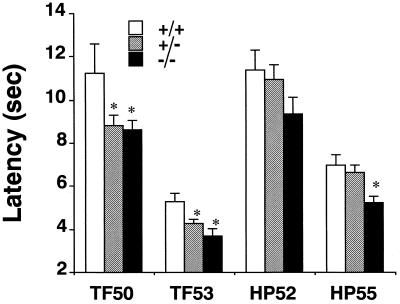
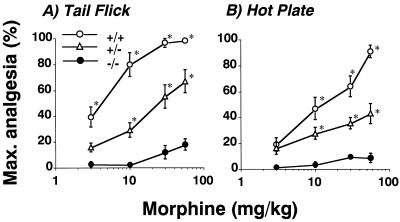
Morphine produces analgesia at opiate receptors expressed in nociceptive circuits. mu, delta, and kappa opiate receptor subtypes are expressed in circuits that can modulate nociception and receive inputs from endogenous opioid neuropeptide ligands. The roles played by each receptor subtype in nociceptive processing in drug-free and morphine-treated states have not been clear, however. We produced homologous, recombinant mu, opiate receptor, heterozygous and homozygous knockout animals that displayed approximately 54% and 0% of wild-type levels of mu receptor expression, respectively. These mice expressed kappa receptors and delta receptors at near wild-type levels. Untreated knockout mice displayed shorter latencies on tail flick and hot plate tests for spinal and supraspinal nociceptive responses than wild-type mice. These findings support a significant role for endogenous opioid-peptide interactions with mu opiate receptors in normal nociceptive processing. Morphine failed to significantly reduce nociceptive responses in hot plate or tail flick tests of homozygous mu receptor knockout mice, and heterozygote mice displayed right and downward shifts in morphine analgesia dose-effect relationships. These results implicate endogenous opioid-peptide actions at mu opiate receptors in several tests of nociceptive responsiveness and support mu receptor mediation of morphine-induced analgesia in tests of spinal and supraspinal analgesia.
Figures
References
-
- Evans C J, Keith D J, Morrison H, Magendzo K, Edwards R H. Science. 1992;258:1952–1955. - PubMed
-
- Eppler C M, Hulmes J D, Wang J B, Johnson B, Corbett M, Luthin D R, Uhl G R, Linden J. J Biol Chem. 1993;268:26447–26451. - PubMed
-
- Chen Y, Mestek A, Liu J, Hurley J A, Yu L. Mol Pharmacol. 1993;44:8–12. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
