

Why do mitochondria synthesize fatty acids? Evidence for involvement in lipoic acid production
- PMID: 9037098
- PMCID: PMC19836
- DOI: 10.1073/pnas.94.4.1591
Why do mitochondria synthesize fatty acids? Evidence for involvement in lipoic acid production
Abstract
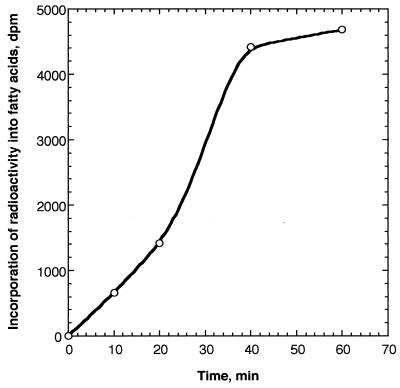
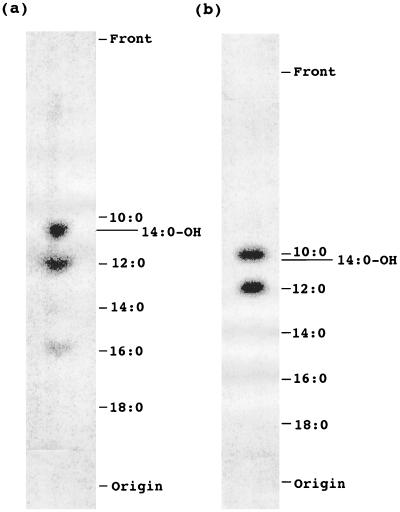
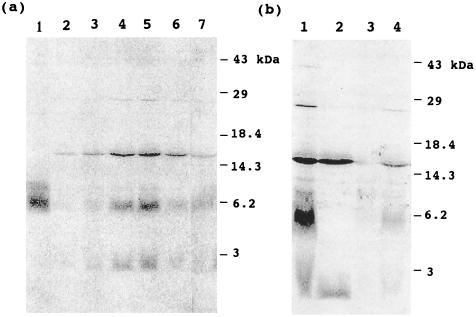
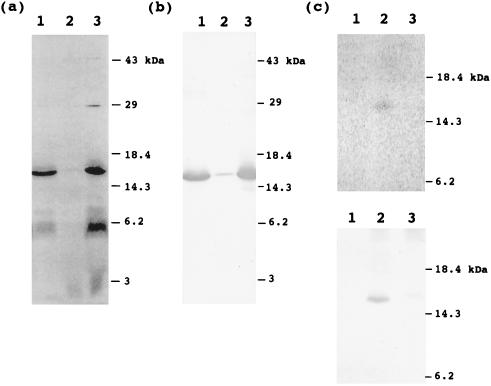
The function of acyl carrier protein (ACP) in mitochondria isolated from pea leaves has been investigated. When pea leaf mitochondria were labeled with [2-14C] malonic acid in vitro, radioactivity was incorporated into fatty acids, and, simultaneously, ACP was acylated. [1-14C]Acetate was much less effective as a precursor for fatty acid synthesis, suggesting that mitochondria do not possess acetyl-CoA carboxylase. The incorporation of radioactivity from [2-14C]malonate into fatty acids and the labeling of ACP were inhibited by cerulenin and required ATP and Mg2+. These findings indicate that plant mitochondria contain not only ACP, but all enzymes required for de novo fatty acid synthesis. Over 30% of the radioactive products from pea mitochondria labeled with [2-14C]malonate were recovered in H protein, which is a subunit of glycine decarboxylase and contains lipoic acid as an essential constituent. In similar experiments, the H protein of Neurospora mitochondria was also labeled by [2-14C]malonate. The labeling of pea H protein was inhibited by addition of cerulenin into the assay medium. Together, these findings indicate that ACP is involved in the de novo synthesis of fatty acids in plant mitochondria and that a major function of this pathway is production of lipoic acid precursors.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
