

The neuroprotective activity of group-II metabotropic glutamate receptors requires new protein synthesis and involves a glial-neuronal signaling
- PMID: 9045718
- PMCID: PMC6793767
- DOI: 10.1523/JNEUROSCI.17-06-01891.1997
The neuroprotective activity of group-II metabotropic glutamate receptors requires new protein synthesis and involves a glial-neuronal signaling
Abstract
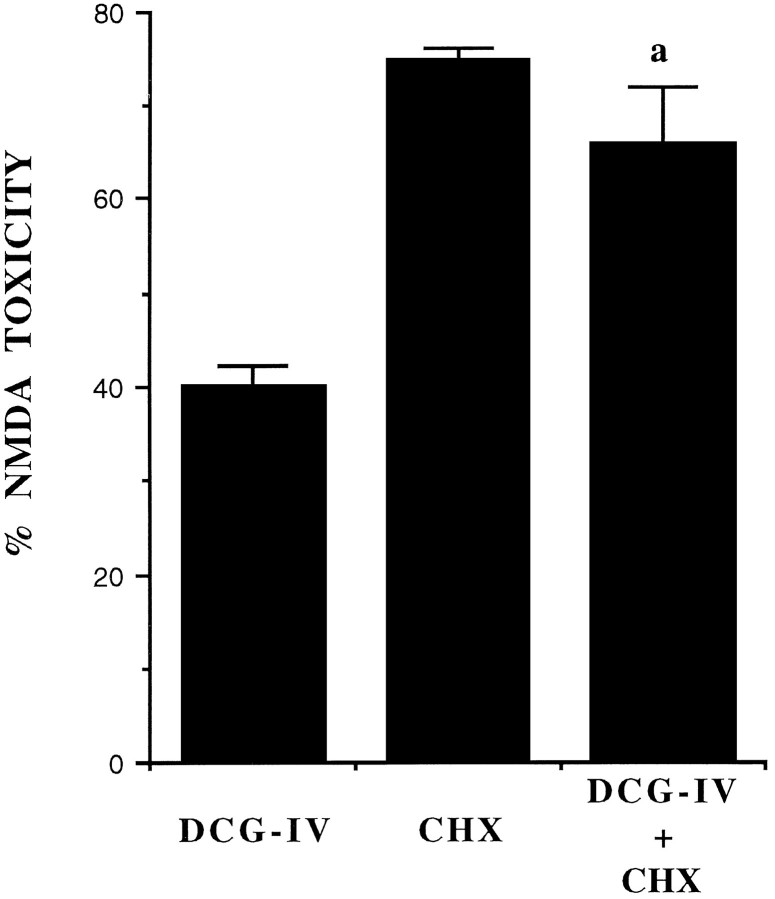
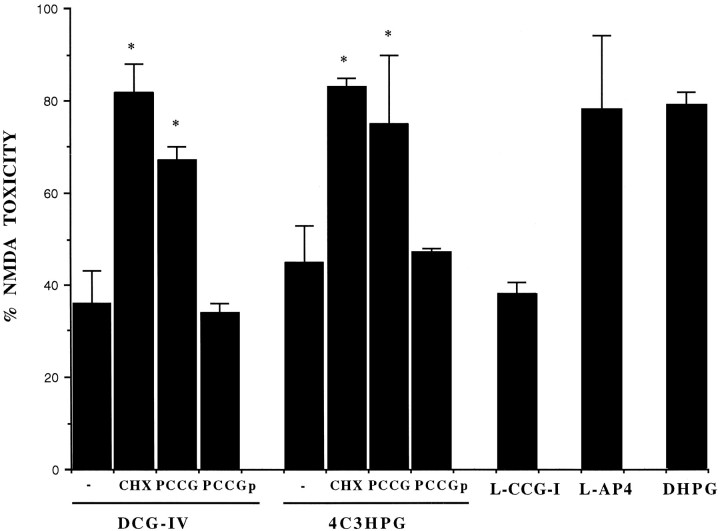
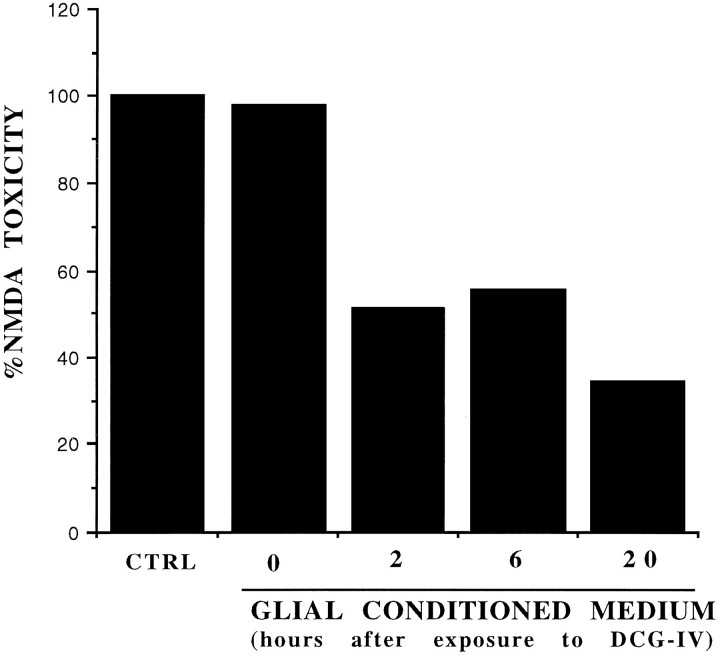
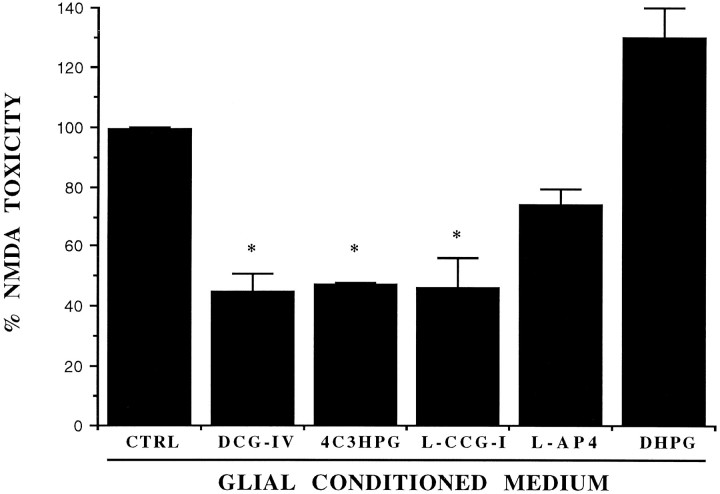
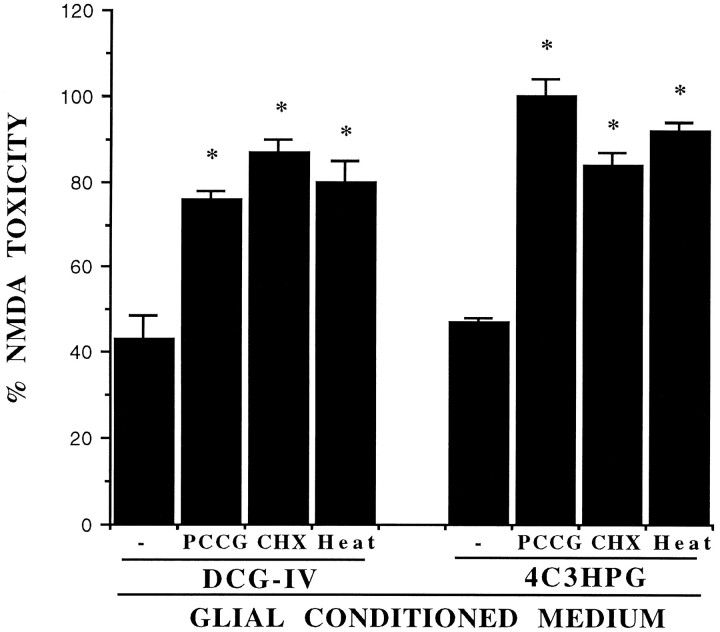
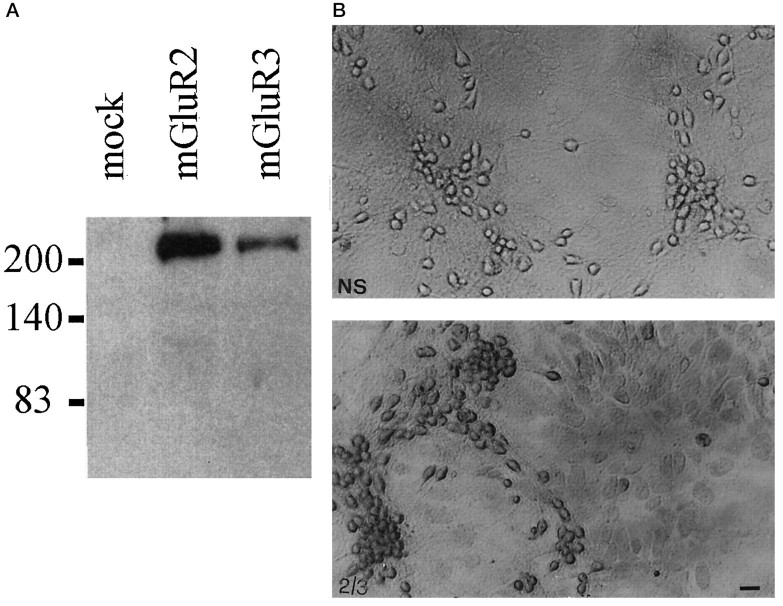
The group-II metabotropic glutamate (mGlu) receptor agonists (2S,1'R, 2'R,3'R)-2-(2,3-dicarboxycyclopropyl)glycine (DCG-IV), S-4-carboxy-3-hydroxyphenylglycine (4C3HPG), and (2S,1'S, 2'S)-2-(carboxycyclopropyl)glycine (L-CCG-I) protected mouse cortical neurons grown in mixed cultures against excitotoxic degeneration induced by a 10 min pulse with NMDA. Protection was observed not only when agonists were added in combination with NMDA but also when they were transiently applied to cultures 6-20 hr before the NMDA pulse. In both cases, neuroprotection was reduced by the group-II mGlu receptor antagonist (2S,1'S,2'S,3'R)-2-(2'-carboxy-3'-phenylcyclopropyl)glycine (PCCG-IV), as well as by the protein synthesis inhibitor cycloheximide (CHX). Both neurons and astrocytes in mixed cultures were immunostained with an antibody that recognized mGlu2 and mGlu3 receptors in recombinant cells. To determine whether astrocytes played any role in the neuroprotection mediated by group-II mGlu receptors, we exposed pure cultures of cortical astrocytes to DCG-IV, 4C3HPG, or L-CCG-I for 10 min. The astrocyte medium collected 2-20 hr after the exposure to any of these drugs was highly neuroprotective when transferred to mixed cultures treated with NMDA. This protective activity was reduced when CHX was applied to astrocyte cultures immediately after the transient exposure to group-II mGlu receptor agonists. We conclude that neuroprotection mediated by group-II mGlu receptors in cultured cortical cells requires new protein synthesis and involves an interaction between neurons and astrocytes.
Figures
References
-
- Altemus KL, Colwell CS, Levine MS. Modulation of excitotoxic damage in the neostriatum by metabotropic glutamate receptors. Soc Neurosci Abstr. 1995;21:217.6.
-
- Balazs R, Miller S, Romano C, de Vries A, Cotman CW. Metabotropic glutamate receptors in cerebral cortical astrocytes: pharmacological properties of mGluR5 and interaction between PLC- and adenylate cyclase (AC)-coupled transduction systems. 2nd International Meeting on Metabotropic Glutamate Receptors. Neuropharmacology. 1996;35:A2.
-
- Bruno V, Copani A, Battaglia G, Raffaele R, Shinozaki H, Nicoletti F. Protective effect of the metabotropic glutamate receptor agonist, DCG-IV, against excitotoxic neuronal death. Eur J Pharmacol. 1994;256:109–112. - PubMed
-
- Bruno V, Battaglia G, Copani A, Giffard RG, Raciti G, Raffaele R, Shinozaki H, Nicoletti F. Activation of class II or III metabotropic glutamate receptors protects cultured cortical neurons against excitotoxic degeneration. Eur J Neurosci. 1995;7:1906–1913. - PubMed
-
- Buisson A, Choi DW. The inhibitory mGluR agonist, S-4-carboxy-3-hydroxy-phenylglycine selectively attenuates NMDA neurotoxicity and oxygen-glucose deprivation-induced neuronal death. Neuropharmacology. 1995;34:1081–1087. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources