

Cell fate specification and symmetrical/asymmetrical divisions in the developing cerebral cortex
- PMID: 9045730
- PMCID: PMC6793772
- DOI: 10.1523/JNEUROSCI.17-06-02018.1997
Cell fate specification and symmetrical/asymmetrical divisions in the developing cerebral cortex
Abstract
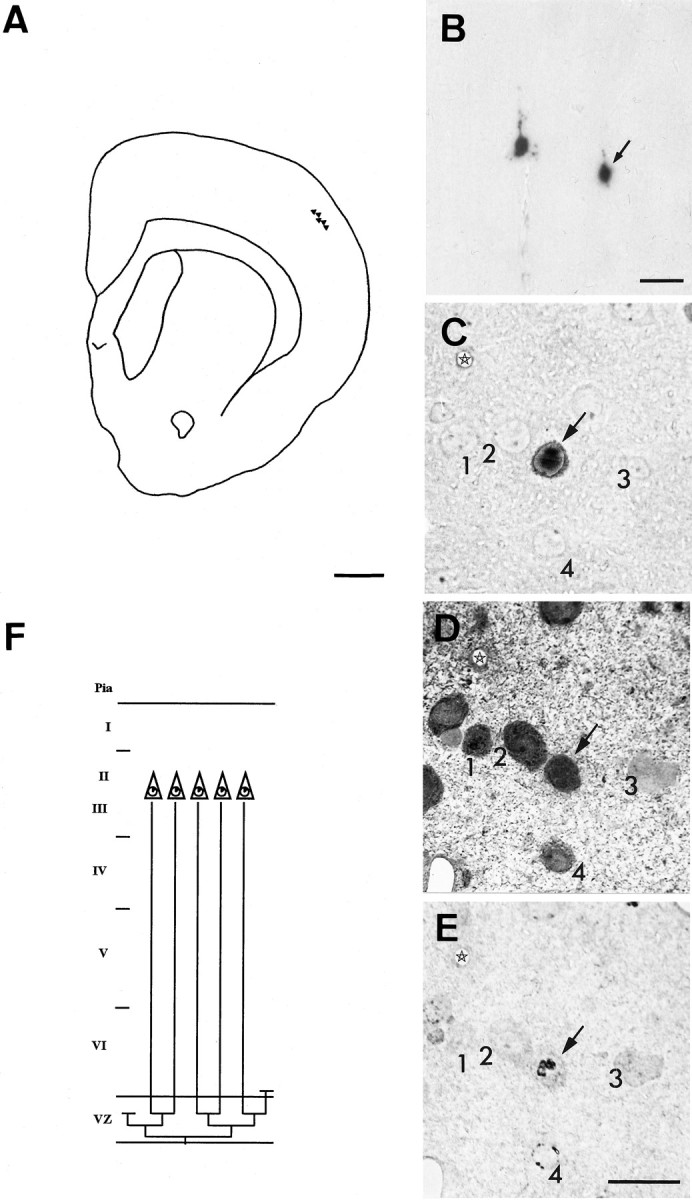
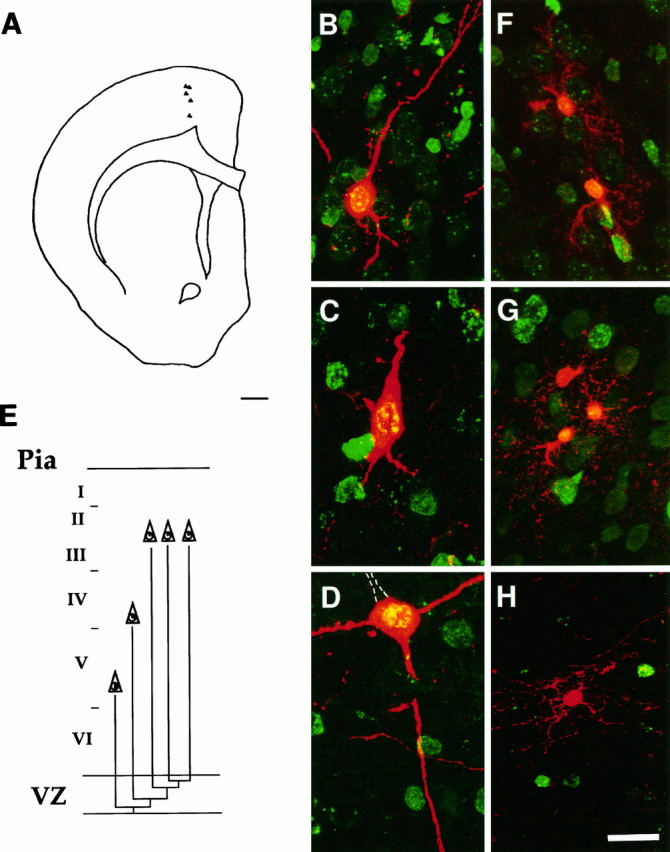
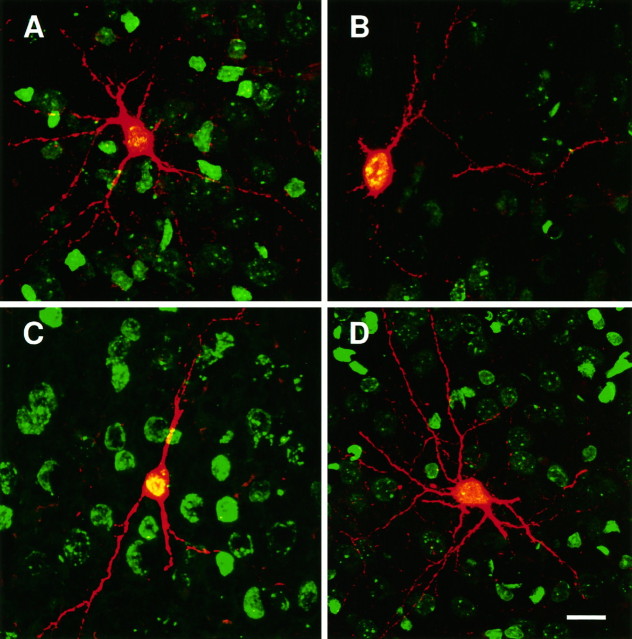
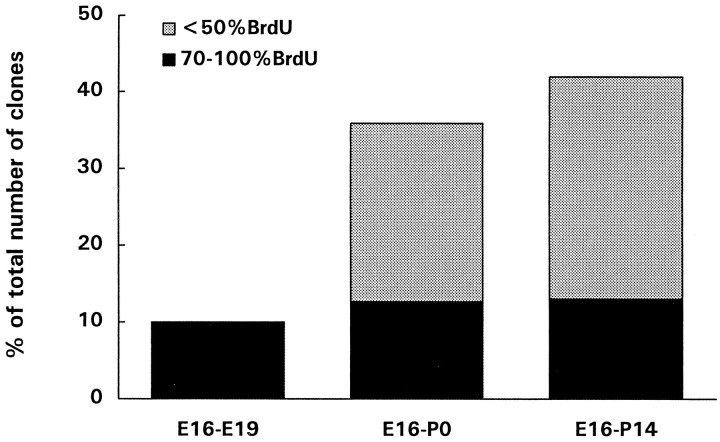
Two different modes of cell division are adopted by progenitor cells to generate the neurons and glia of the cerebral cortex: they either divide symmetrically to generate other progenitors or a pair of postmitotic cells or divide asymmetrically to generate both a progenitor and a postmitotic cell. In this study we used a lineage marker, the BAG retrovirus, in embryonic day 16 rats in combination with bromodeoxyuridine (BrdU) to identify patterns of cell generation in the cerebral cortex, and investigated the relationship between the phenotype of cells and the history of their lineages. The location, phenotype and birth order of clonally related cells were studied in the subsequent 3 weeks. Only pyramidal neurons and/or astrocytes formed discrete clusters in which several generations of family members were present, whereas nonpyramidal neurons were found exclusively in pairs or as single cells. Analysis of BrdU levels in these cells showed that nonpyramidal neurons were originally part of larger clones and were found dispersed in the neocortex because of tangential migration of their progenitors, dispersion of postmitotic cells, or death of clonal relatives. These results suggest that both symmetrical and asymmetrical division can be adopted by progenitor cells to generate cortical neurons and glial cells and that cell extrinsic events contribute to the isolation of nonpyramidal neurons.
Figures

References
-
- Acklin SE, van der Kooy D. Clonal heterogeneity in the germinal zone of the developing rat telencephalon. Development. 1993;118:175–192. - PubMed
-
- Anderson DJ. The neural crest lineage problem: neuropoiesis? Neuron. 1989;3:1–12. - PubMed
-
- Artavanis-Tsakonas S, Matsuno K, Fortini ME. Notch signaling. Science. 1995;268:225–232. - PubMed
-
- Austin CP, Cepko CL. Cellular migration patterns in the developing mouse cerebral cortex. Development. 1990;110:713–732. - PubMed
-
- Bayer SA, Altman J. Neocortical development. Raven; New York: 1991.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources