

Local homogeneity of cell cycle length in developing mouse cortex
- PMID: 9045735
- PMCID: PMC6793765
- DOI: 10.1523/JNEUROSCI.17-06-02079.1997
Local homogeneity of cell cycle length in developing mouse cortex
Abstract
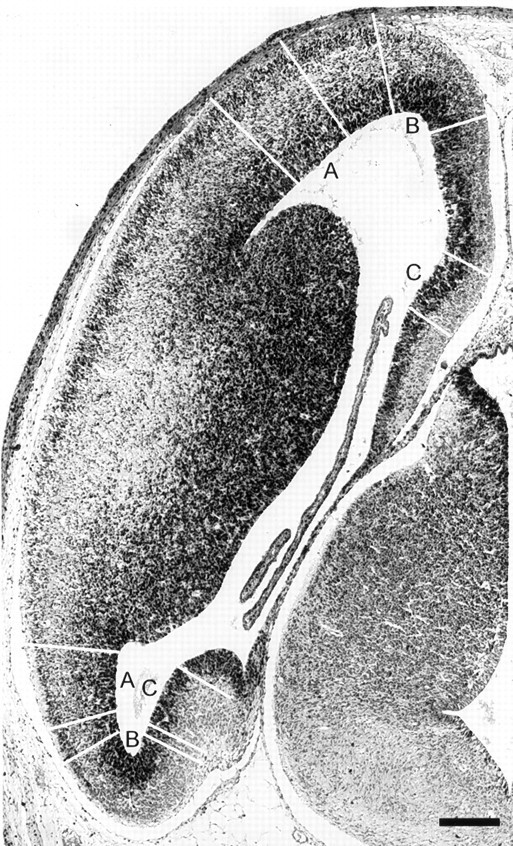
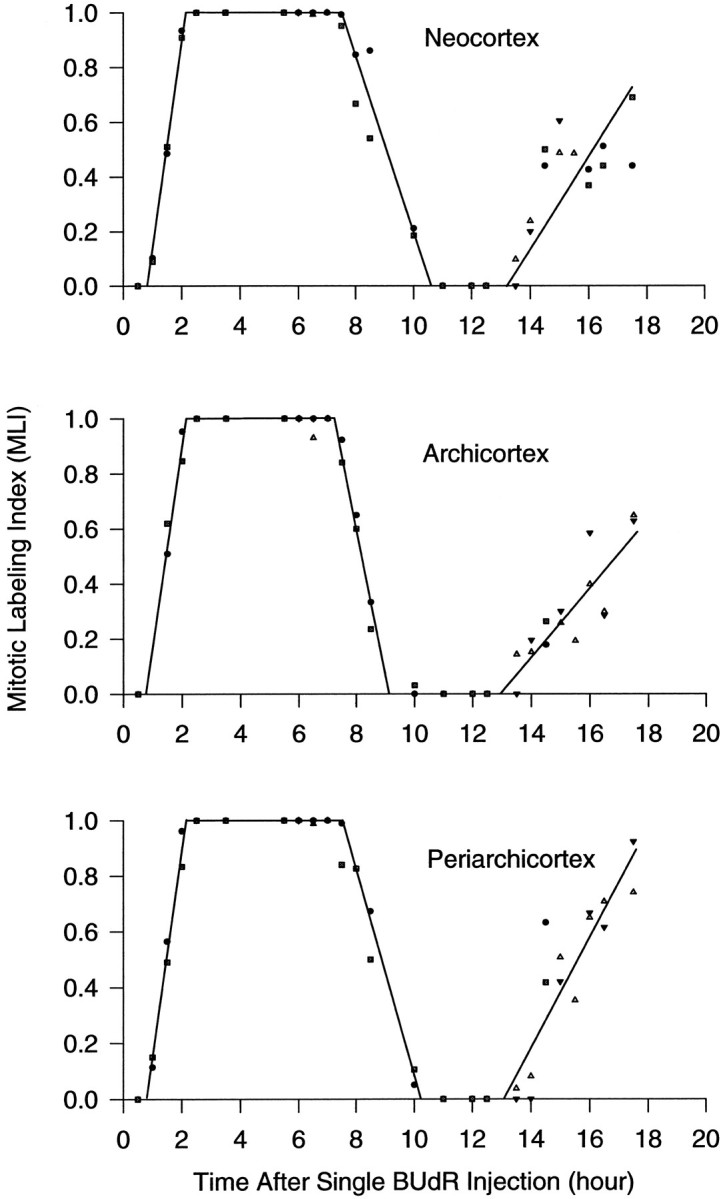
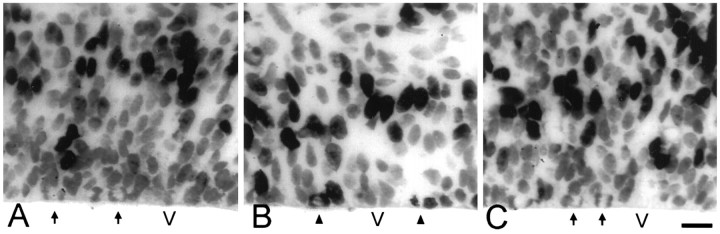
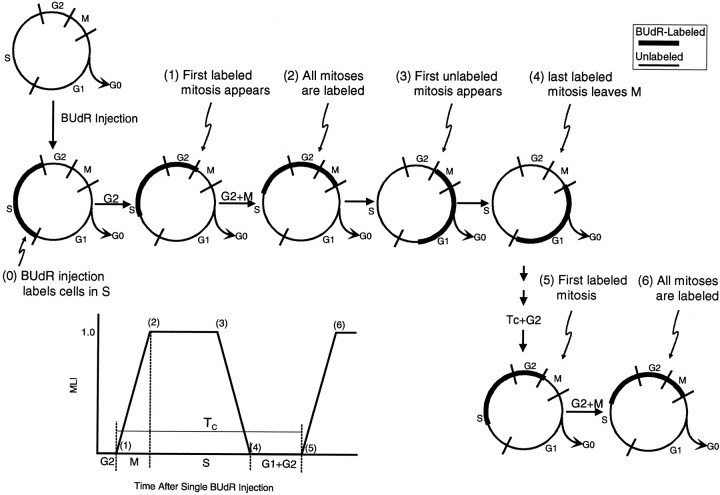
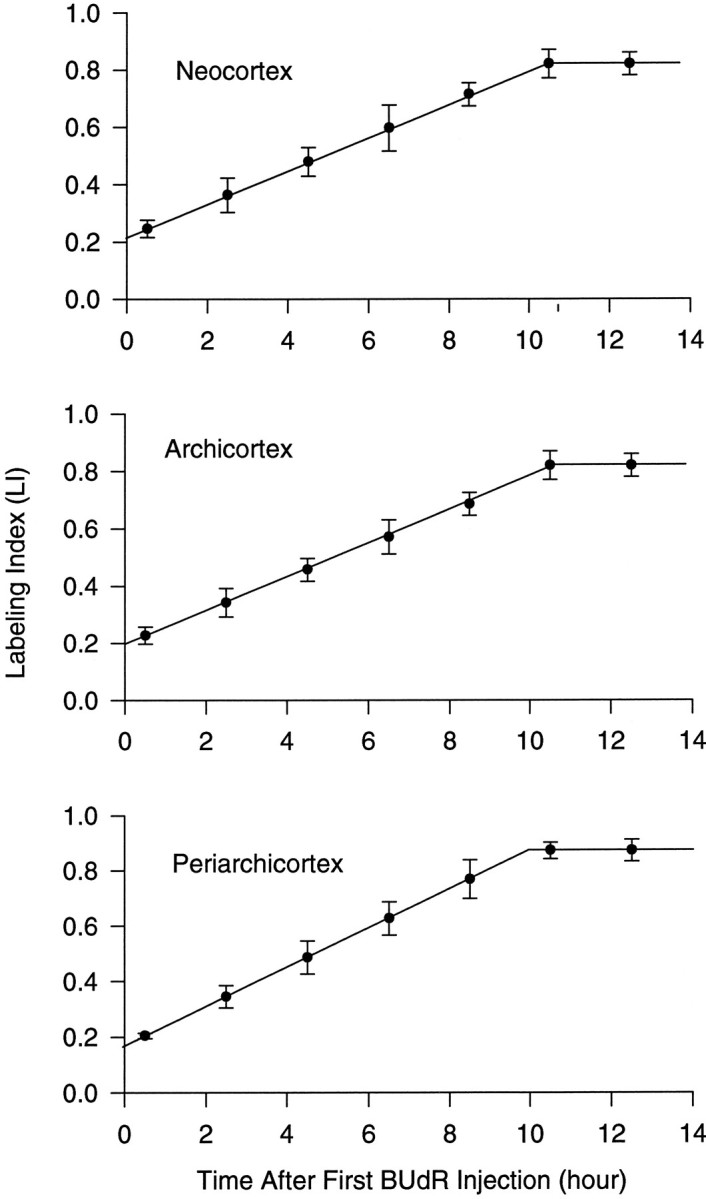
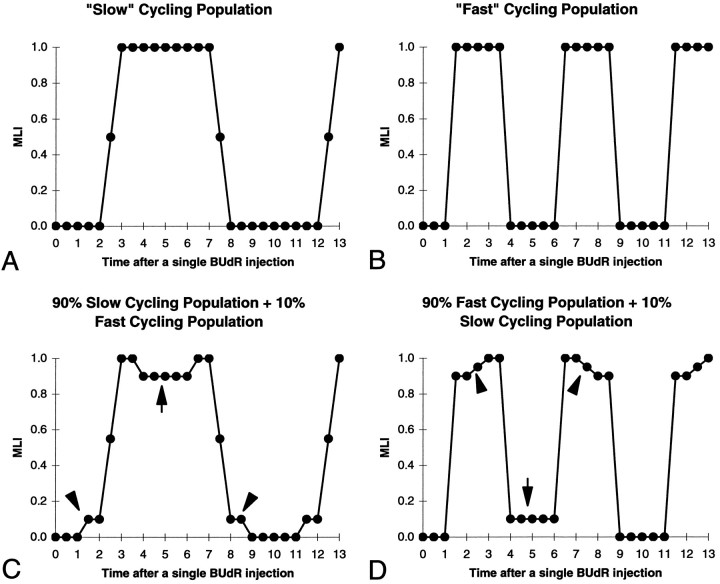
We have measured the amount of variation in the length of the cell cycle for cells in the pseudostratified ventricular epithelium (PVE) of the developing cortex of mice on embryonic day 14. Our measurements were made in three cortical regions (i.e., the neocortex, archicortex, and periarchicortex) using three different methods: the cumulative labeling method (CLM), the percent labeled mitoses (PLM) method, and a comparison of the time needed for the PLM to ascend from 0 to 100% with the time needed for the PLM to descend from 100 to 0%. These 3 different techniques provide different perspectives on the cytokinetic parameters. Theoretically, CLM gives an estimate for a maximum value of the total length of the cell cycle (TC), whereas PLM gives an estimate of a minimum value of TC. The difference between these two estimates indicates that the range for TC is +/-1% of the mean TC for periarchicortex, +/-7% for neocortex, and +/-8% for archicortex. This was confirmed by a lengthening of the PLM descent time in comparison with its ascent time. The sharpness of the transitions and the flatness of the plateau of the PLM curves indicate that 99% of the proliferating cells are within this narrow estimated range for TC; hence, only approximately 1% deviate outside of a relatively restricted range from the average TC of the population. In the context of the possible existence within the cortical PVE of two populations with markedly dissimilar cell cycle kinetics from the mean, one such population must comprise approximately 99% of the total population, and the other, if it exists, is only approximately 1% of the total. This seems to be true for all three cortical regions. The narrow range of TC indicates a homogeneity in the cell cycle length for proliferating cells in three different cortical regions, despite the fact that progenitor cells of different lineages may be present. It further predicts the existence of almost synchronous interkinetic nuclear movements of the proliferating cells in the ventricular zone during early development of the cerebral cortex.
Figures
References
-
- Angevine JB, Jr, Sidman RL. Autoradiographic study of cell migration during histogenesis of cerebral cortex in the mouse. Nature. 1961;192:766–768. - PubMed
-
- Cai L, Hayes NL, Nowakowski RS. Comparison of the cumulative S-phase labeling method and the percent labeled mitoses method in the developing cerebral cortex. Soc Neurosci Abstr. 1993;19:30.
-
- Caviness VS., Jr Neocortical histogenesis in normal and reeler mice: a developmental study based upon [3H]thymidine autoradiography. Dev Brain Res. 1982;4:293–302. - PubMed
-
- Caviness VS, Jr, Sidman RL. Time of origin of corresponding cell classes in the cerebral cortex of normal and reeler mutant mice: an autoradiographic analysis. J Comp Neurol. 1973;148:141–151. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources