

Identification of a novel stage of ribosome/nascent chain association with the endoplasmic reticulum membrane
- PMID: 9087438
- PMCID: PMC2132505
- DOI: 10.1083/jcb.136.6.1213
Identification of a novel stage of ribosome/nascent chain association with the endoplasmic reticulum membrane
Abstract
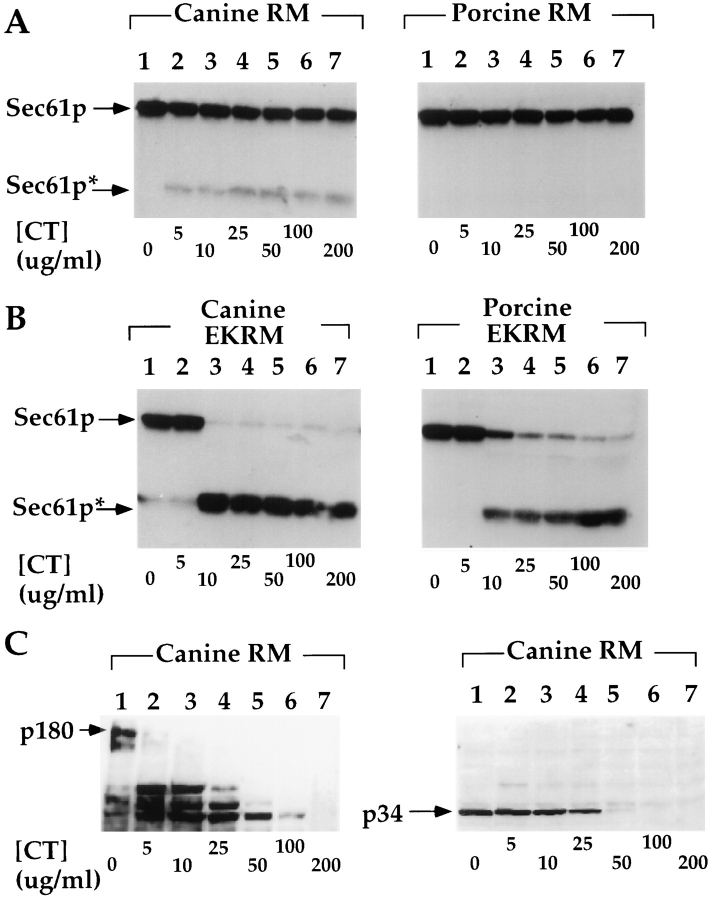
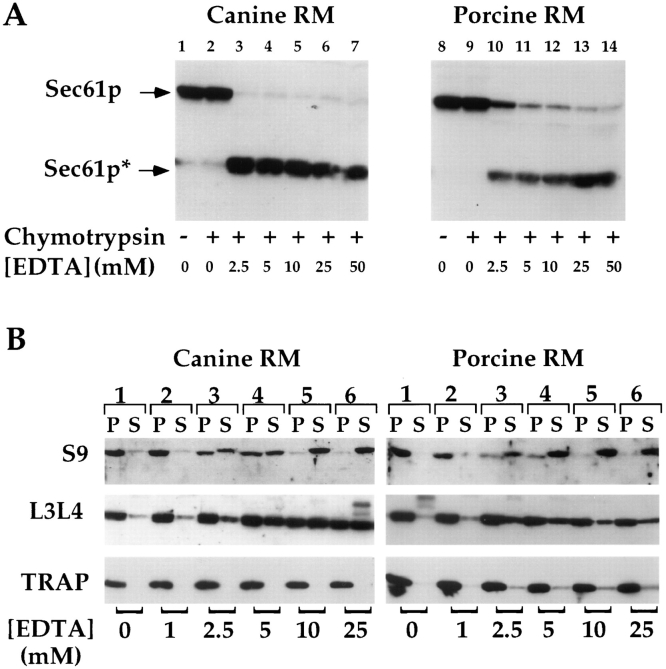
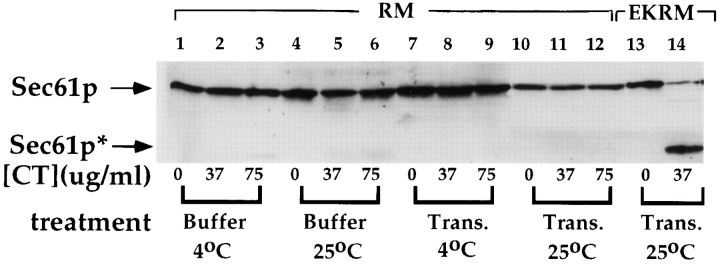
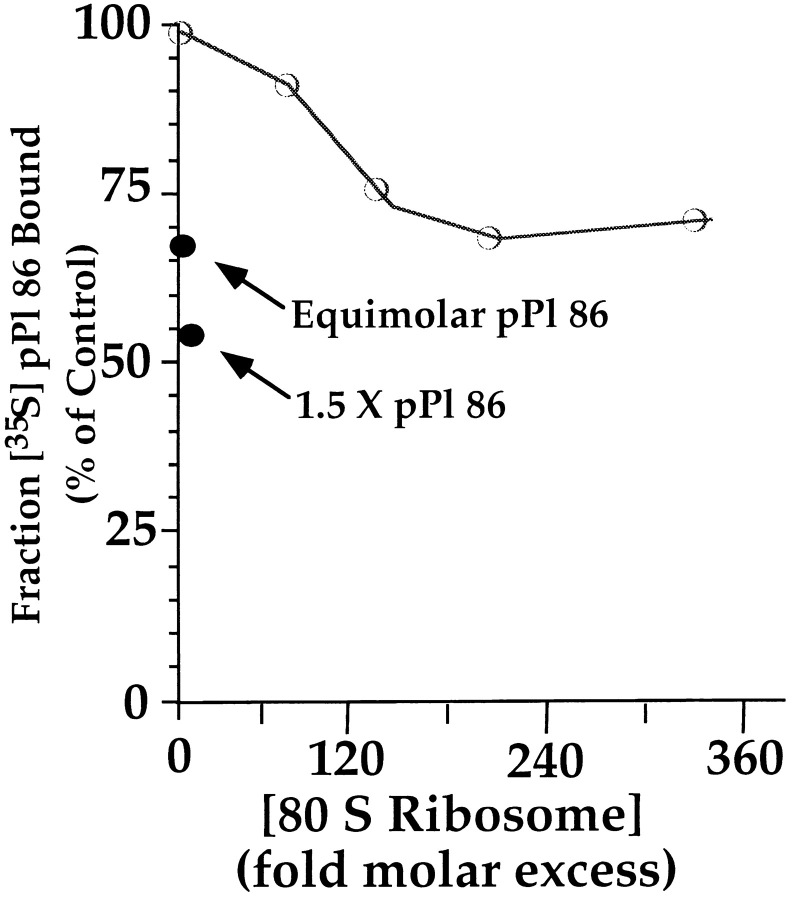
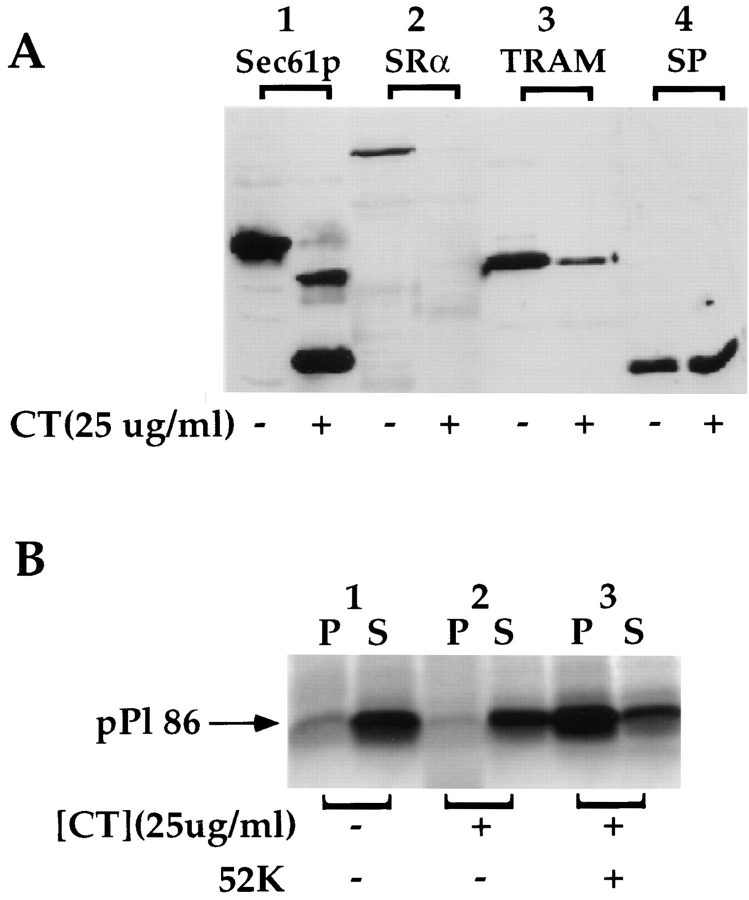
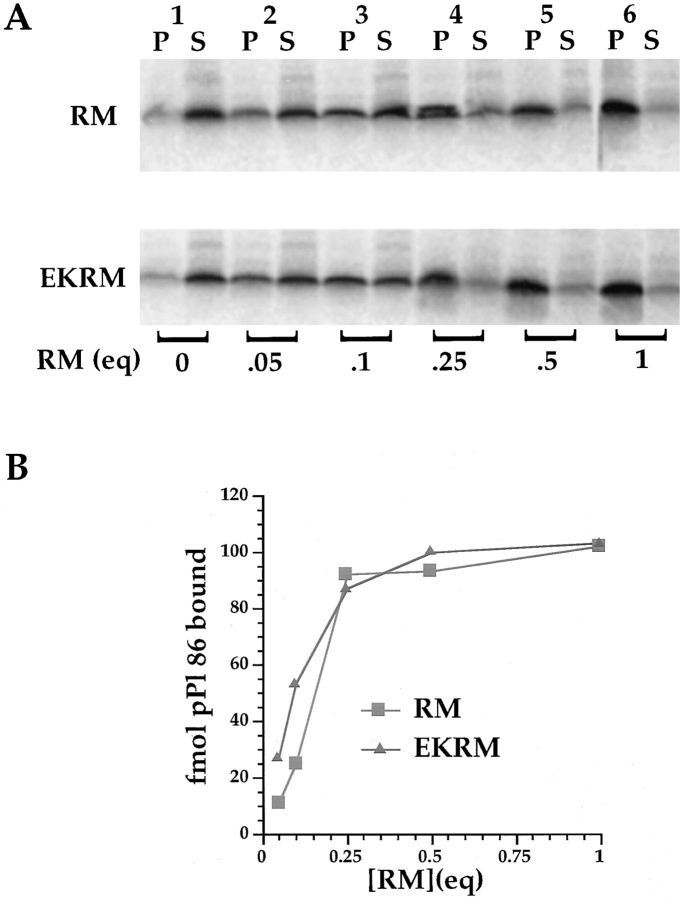
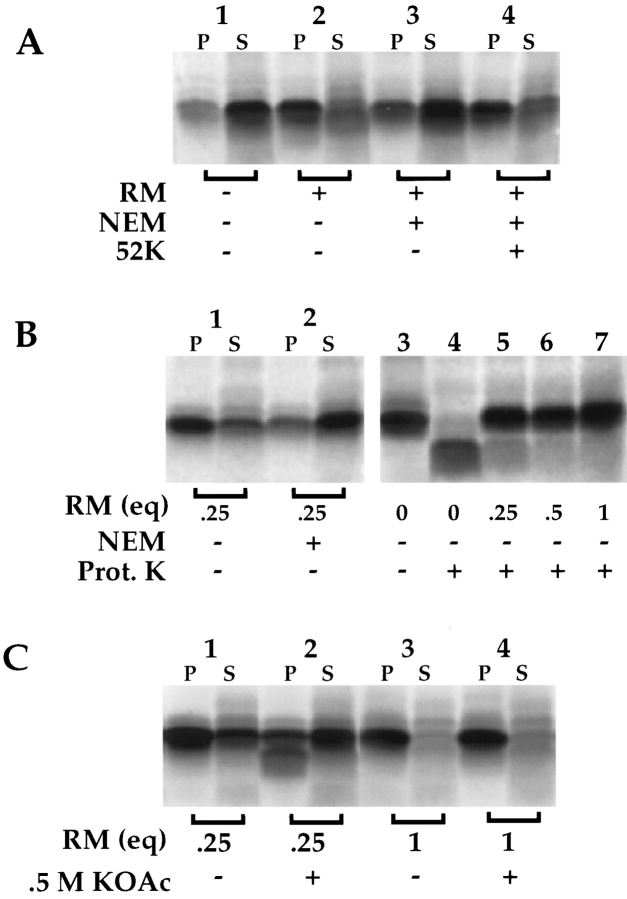
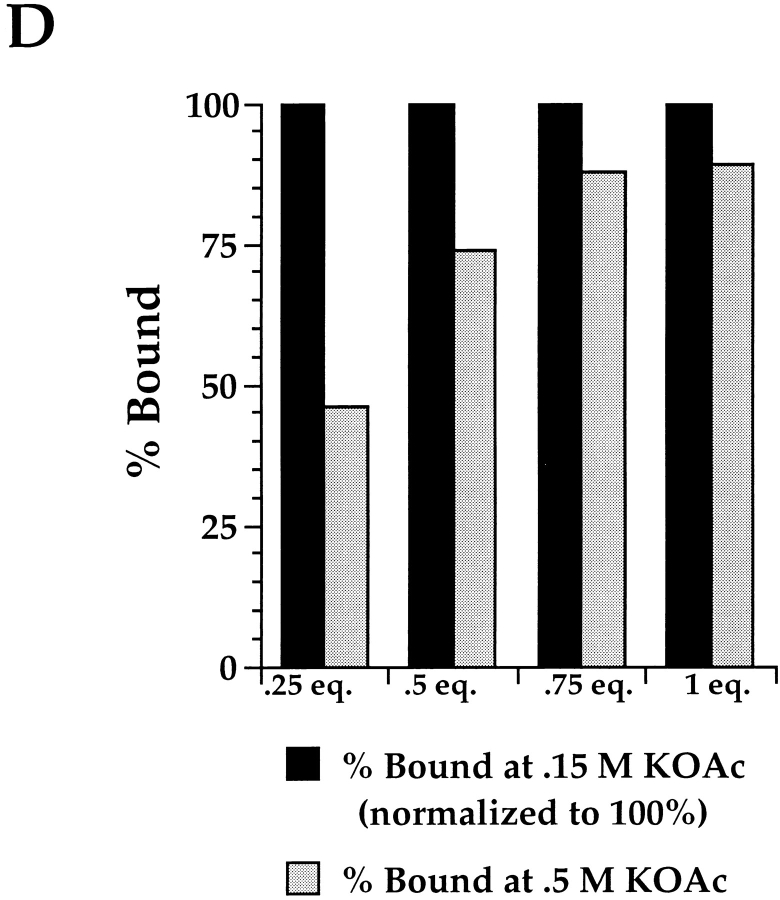
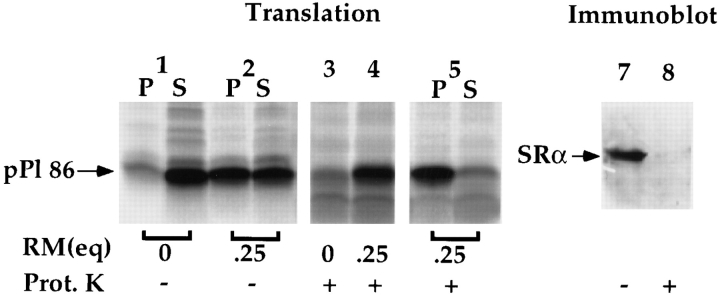
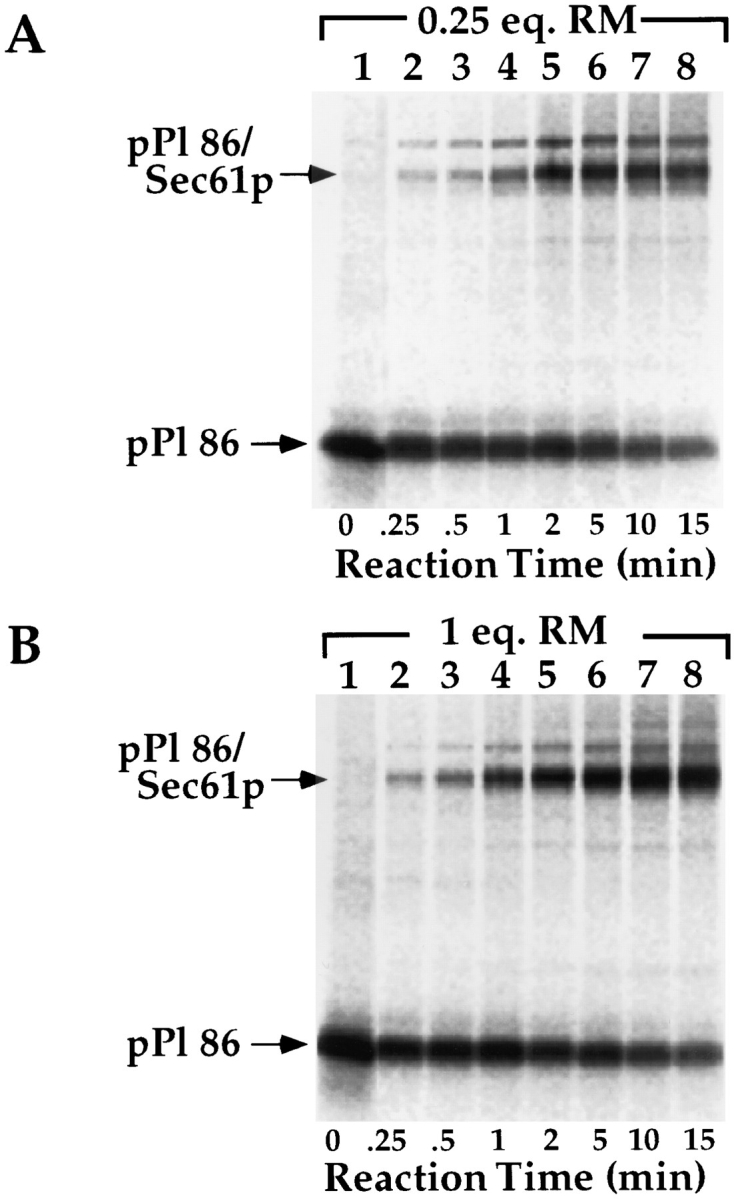
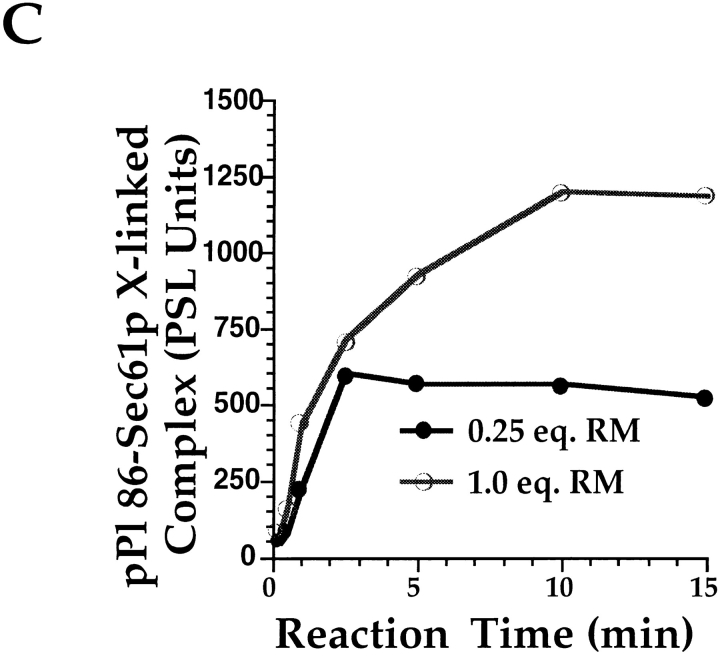
Protein translocation in the mammalian endoplasmic reticulum (ER) occurs cotranslationally and requires the binding of translationally active ribosomes to components of the ER membrane. Three candidate ribosome receptors, p180, p34, and Sec61p, have been identified in binding studies with inactive ribosomes, suggesting that ribosome binding is mediated through a receptor-ligand interaction. To determine if the binding of nascent chain-bearing ribosomes is regulated in a manner similar to inactive ribosomes, we have investigated the ribosome/nascent chain binding event that accompanies targeting. In agreement with previous reports, indicating that Sec61p displays the majority of the ER ribosome binding activity, we observed that Sec61p is shielded from proteolytic digestion by native, bound ribosomes. The binding of active, nascent chain bearing ribosomes to the ER membrane is, however, insensitive to the ribosome occupancy state of Sec61p. To determine if additional, Sec61p independent, stages of the ribosome binding reaction could be identified, ribosome/nascent chain binding was assayed as a function of RM concentration. At limiting RM concentrations, a protease resistant ribosome-membrane junction was formed, yet the nascent chain was salt extractable and cross-linked to Sec61p with low efficiency. At nonlimiting RM concentrations, bound nascent chains were protease and salt resistant and cross-linked to Sec61p with higher efficiency. On the basis of these and other data, we propose that ribosome binding to the ER membrane is a multi-stage process comprised of an initial, Sec61p independent binding event, which precedes association of the ribosome/nascent chain complex with Sec61p.
Figures
References
-
- Bacher G, Lutcke H, Jungnickel B, Rapoport TA, Dobberstein B. Regulation by the ribosome of the GTPase of the signal recognition particle during protein targeting. Nature (Lond) 1996;381:248–251. - PubMed
-
- Borgese D, Blobel G, Sabatini DD. In vitro exchange of ribosomal subunits between free and membrane-bound ribosomes. J Mol Biol. 1973;74:415–438. - PubMed
-
- Borgese N, Mok W, Kreibich G, Sabatini DD. Ribosomal-membrane interaction: in vitro binding of ribosomes to microsomal membranes. J Mol Biol. 1974;88:559–580. - PubMed
