

beta-Actin messenger RNA localization and protein synthesis augment cell motility
- PMID: 9087442
- PMCID: PMC2132521
- DOI: 10.1083/jcb.136.6.1263
beta-Actin messenger RNA localization and protein synthesis augment cell motility
Erratum in
- J Cell Biol 1997 Jun 30;137(7):1683
Abstract
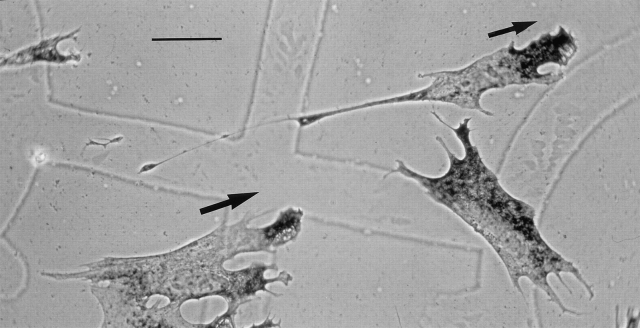
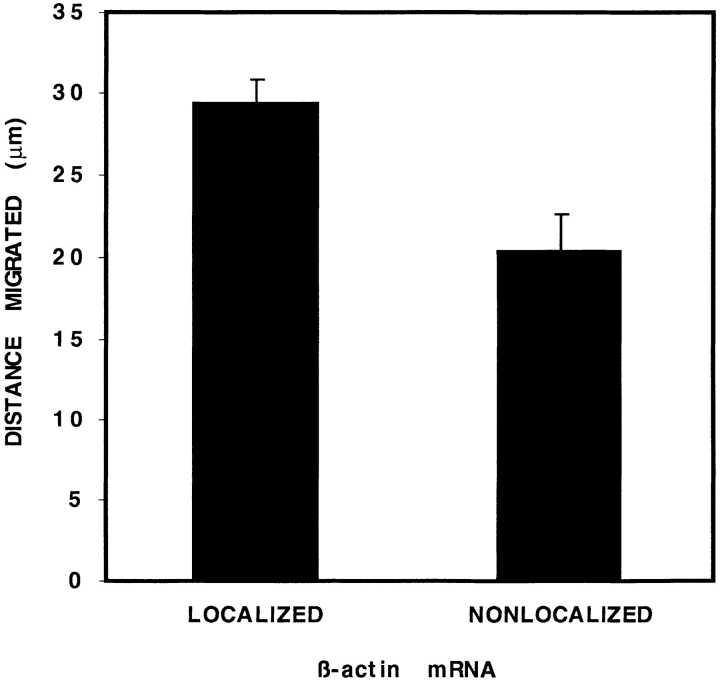
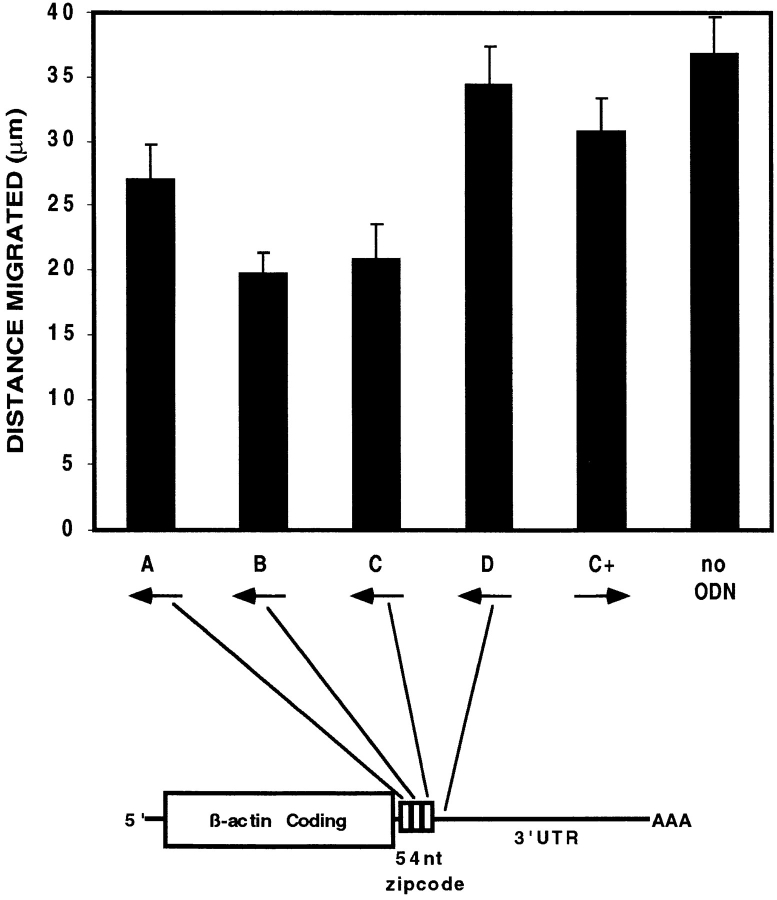
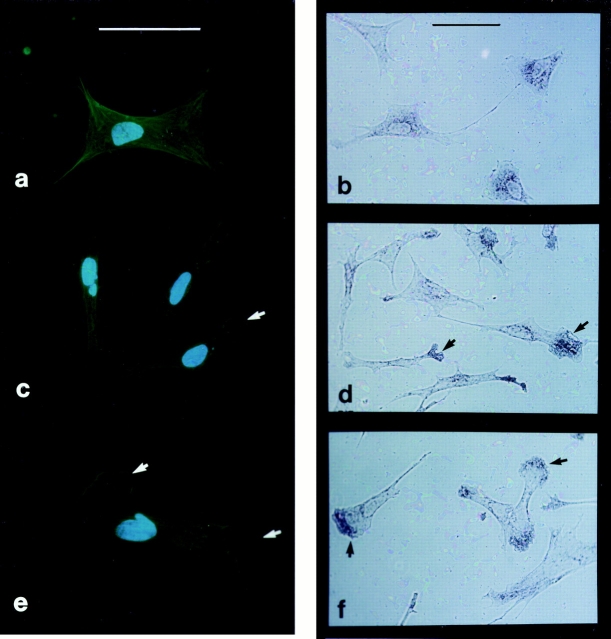
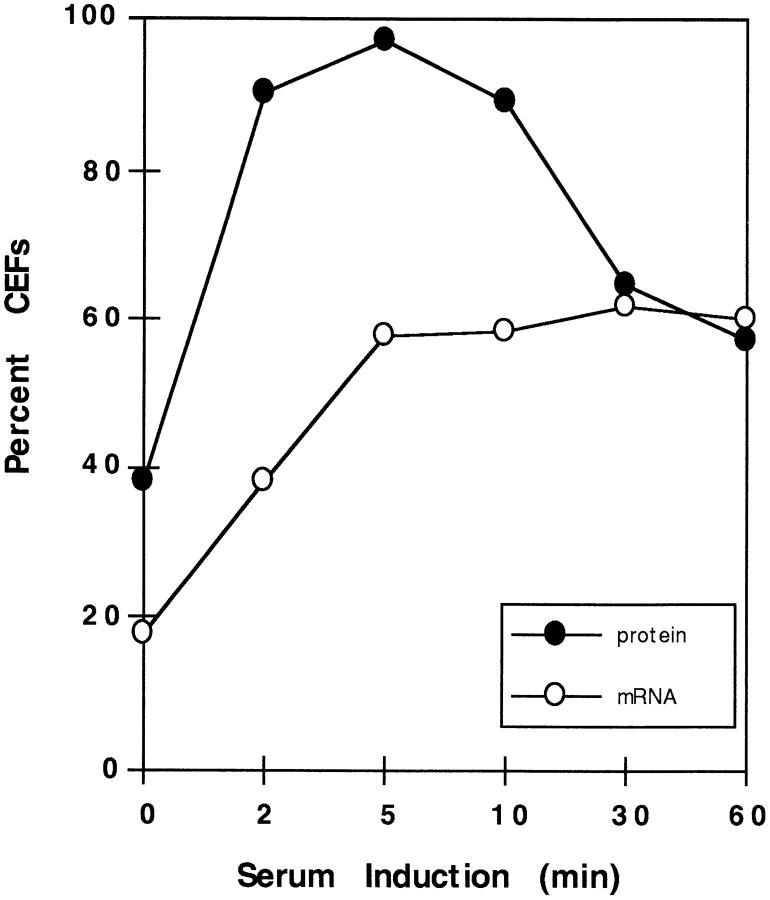
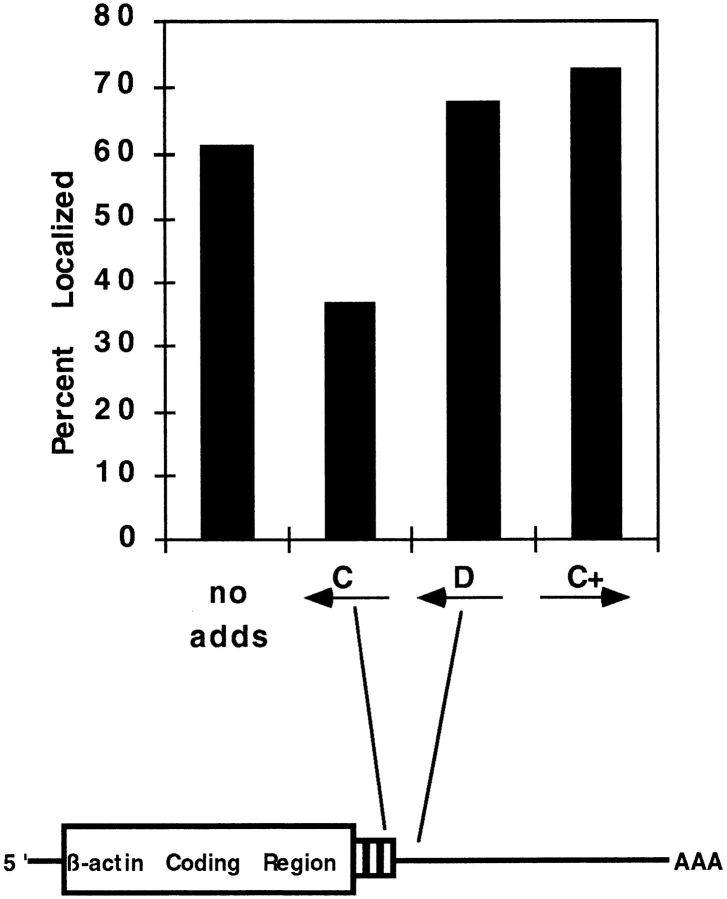
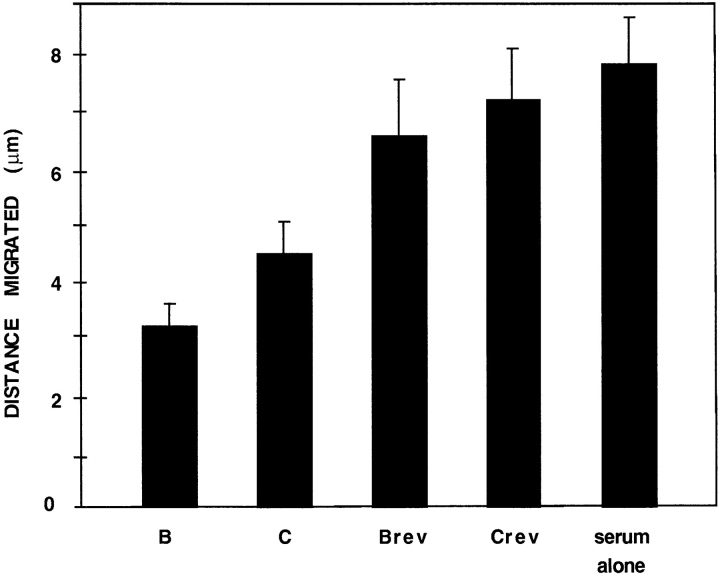
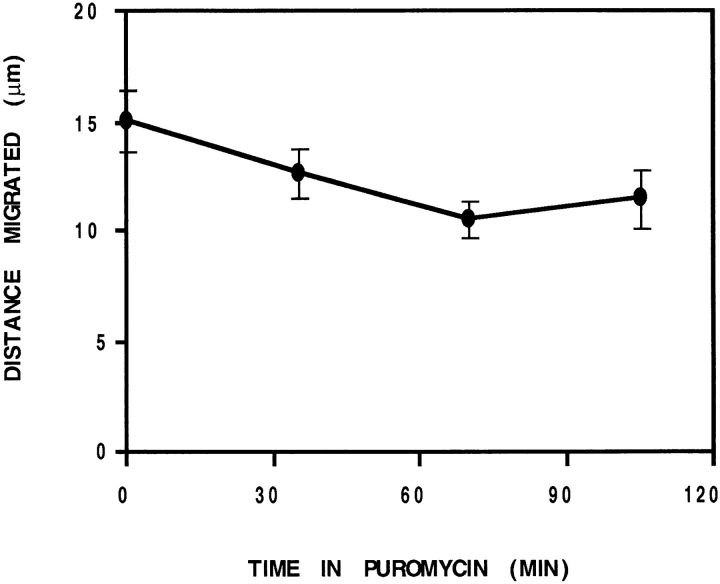
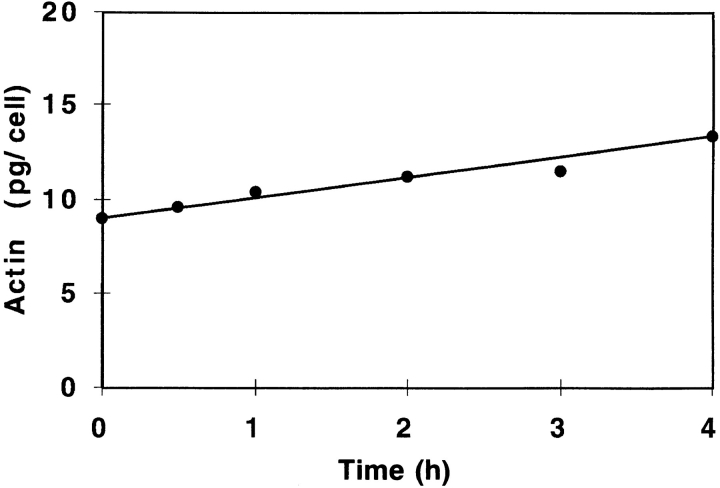
In chicken embryo fibroblasts (CEFs), beta-actin mRNA localizes near an actin-rich region of cytoplasm specialized for motility, the lamellipodia. This localization is mediated by isoform-specific 3'-untranslated sequences (zipcodes) and can be inhibited by antizipcode oligodeoxynucleotides (ODNs) (Kislauskis, E.H., X.-C. Zhu, and R.H. Singer. 1994. J. Cell Biol. 127: 441-451). This inhibition of beta-actin mRNA localization resulted in the disruption of fibroblast polarity and, presumably, cell motility. To investigate the role of beta-actin mRNA in motility, we correlated time-lapse images of moving CEFs with the distribution of beta-actin mRNA in these cells. CEFs with localized beta-actin mRNA moved significantly further over the same time period than did CEFs with nonlocalized mRNA. Antizipcode ODN treatment reduced this cell translocation while control ODN treatments showed no effect. The temporal relationship of beta-actin mRNA localization to cell translocation was investigated using serum addition to serum-deprived cultures. beta-actin mRNA was not localized in serum-deprived cells but became localized within minutes after serum addition (Latham, V.M., E.H. Kislauskis, R.H. Singer, and A.F. Ross. 1994. J. Cell Biol. 126:1211-1219). Cell translocation increased over the next 90 min, and actin synthesis likewise increased. Puromycin reduced this cell translocation and blocked this induction in cytosolic actin content. The serum induction of cell movement was also inhibited by antizipcode ODNs. These observations support the hypothesis that beta-actin mRNA localization and consequent protein synthesis augment cell motility.
Figures
References
-
- Bershadsky AD, Gluck U, Denisenko ON, Sklyarova TV, Spector I, Ben-Ze'ev A. The state of actin assembly regulates actin and vinculin expression by a feedback loop. J Cell Sci. 1995;108:1183–1193. - PubMed
-
- Condeelis J. Life at the leading edge: the formation of cell protrusions. Annu Rev Cell Biol. 1993;9:411–444. - PubMed
-
- Darnell, J.E., H. Lodish, D. Baltimore, A. Berk, S.L. Zipursky, and P. Matsudaira. 1995. Molecular Cell Biology. W. H. Freeman, editor. Scientific American Books, Inc., New York. 129.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
