

A functional telomerase RNA swap in vivo reveals the importance of nontemplate RNA domains
- PMID: 9096304
- PMCID: PMC20280
- DOI: 10.1073/pnas.94.7.2823
A functional telomerase RNA swap in vivo reveals the importance of nontemplate RNA domains
Abstract
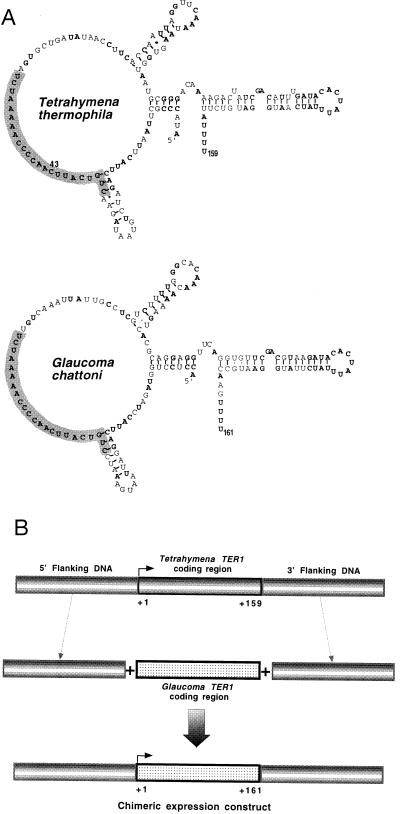
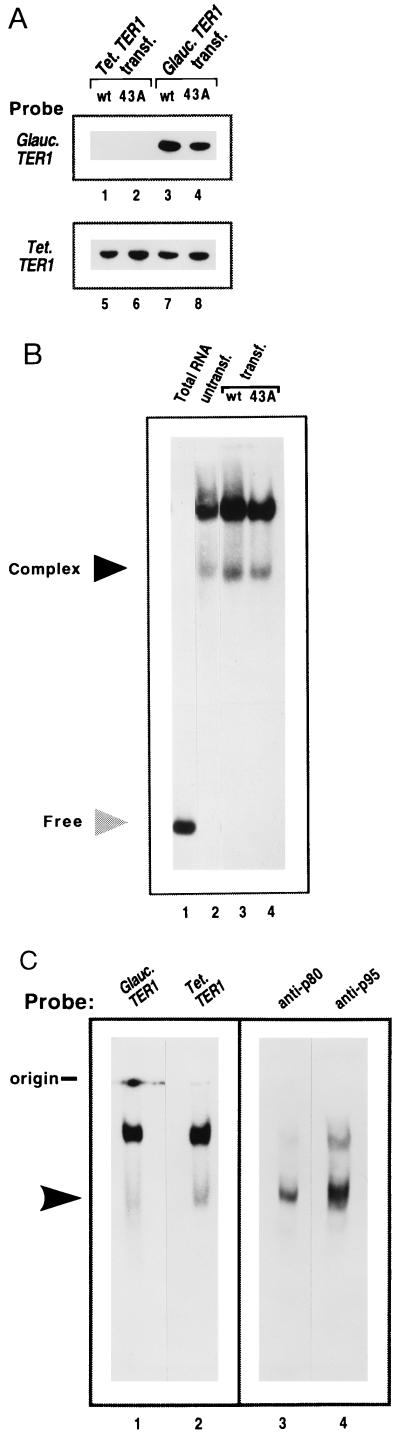
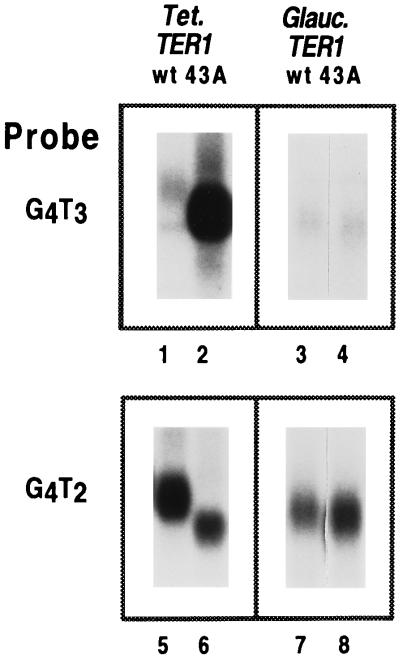
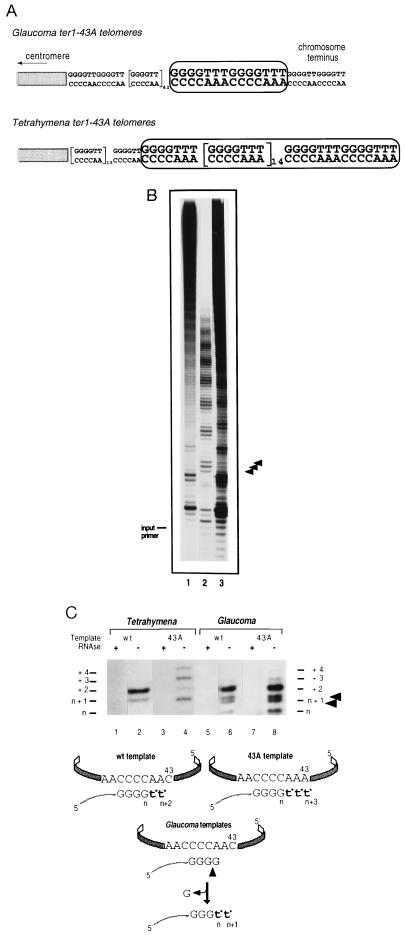
The ribonucleoprotein (RNP) enzyme telomerase is required for replication of eukaryotic chromosomal termini. The RNA moiety of telomerase is essential for enzyme function and provides the template for telomeric DNA synthesis. However, the roles of its nontemplate domains have not been explored. Here we demonstrate that a novel interspecies telomerase RNA swap in vivo creates a functional but aberrant telomerase. Telomerase RNA from the ciliate Glaucoma chattoni was expressed in Tetrahymena thermophila cells. The telomerase RNAs from these two species have almost superimposable secondary structures. The template region base sequence is identical in the two RNAs, but elsewhere their sequences differ by 49%. This hybrid telomerase RNP was enzymatically active but added only short stretches of telomeric repeat tracts in vivo and in vitro. This new enzyme also had a strong, aberrant DNA cleavage activity in vitro. Thus, molecular interactions in the RNP involving nontemplate RNA domains affect specific aspects of telomerase enzyme function, raising the possibility that they may regulate telomerase activity.
Figures
References
-
- Telomeres (1995) eds. Greider C. & Blackburn E. H. (Cold Spring Harbor Lab. Press, Plainview, NY).
-
- Kirk, K. E., Harmon, B. P., Riechardt, I. K, Sedat, J. & Blackburn, E. H. (1997) Science, in press. - PubMed
-
- Greider C W, Blackburn E H. Nature (London) 1989;337:331–337. - PubMed
-
- Gilley D, Lee M S, Blackburn E H. Genes Dev. 1995;9:2214–2226. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
