

Identification of a novel pathway important for proliferation and differentiation of primary erythroid progenitors
- PMID: 9096338
- PMCID: PMC20314
- DOI: 10.1073/pnas.94.7.3016
Identification of a novel pathway important for proliferation and differentiation of primary erythroid progenitors
Abstract
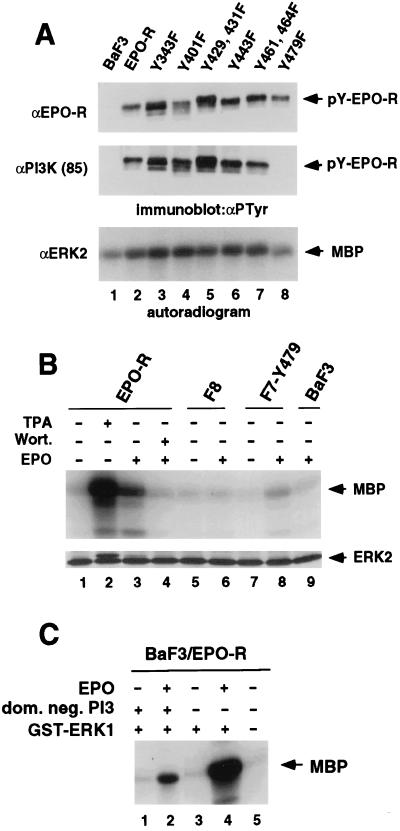
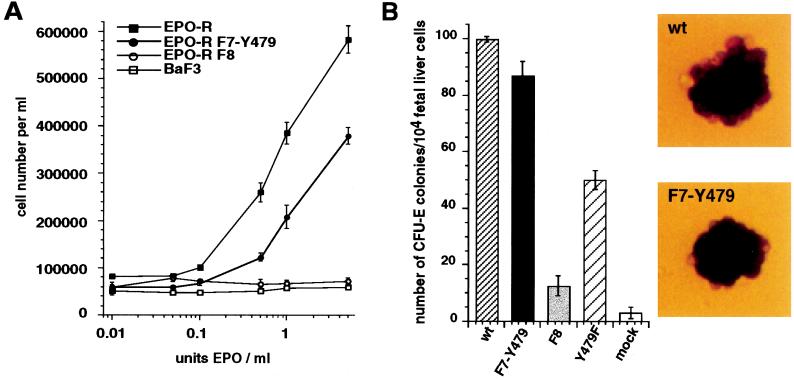
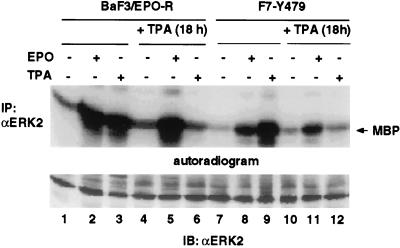
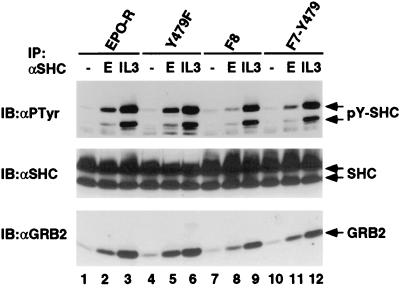
Homodimerization of the erythropoietin (EPO) receptor (EPO-R) in response to EPO binding transiently activates the receptor-associated protein tyrosine kinase JAK2. Tyrosine phosphorylation of the EPO-R creates "docking sites" for SH2 domain(s) in signaling molecules such as the protein tyrosine phosphatases SH-PTP1 and SH-PTP2, phosphoinositide 3-kinase (PI3 kinase), and STAT5. However, little is known about the specific intracellular signals essential for proliferation and differentiation of erythroid progenitors. Here we show that an EPO-R containing only one cytosolic (phospho)tyrosine residue, Y479, induces a signal transduction pathway sufficient for proliferation and differentiation of fetal liver progenitors of erythroid colony-forming units from EPO-R(-/-) mice as well as for proliferation of cultured hematopoietic cells. This cascade involves sequential EPO-induced recruitment of PI3 kinase to the EPO-R and activation of mitogen-activated protein kinase activity, independent of the Shc/Grb2-adapter pathway and of STAT5. Protein kinase C epsilon may be one of the mediators connecting PI3 kinase with the mitogen-activated protein kinase signaling cascade. Our results identify a signaling cascade important in vivo for erythroid cell proliferation and differentiation.
Figures
References
-
- D’Andrea A D, Lodish H F, Wong G G. Cell. 1989;57:277–285. - PubMed
-
- Wrightom N C, Farrell F X, Chang R, Kashyap A K, Barbone F P, Mulcahy L S, Johnson D L, Barrett R W, Jolliffe L K, Dower W J. Science. 1996;273:397–402. - PubMed
-
- Witthuhn B A, Quelle F W, Silvennoinen O, Yi T, Tang B, Miura O, Ihle J N. Cell. 1993;74:227–236. - PubMed
-
- Klingmüller U, Lorenz U, Cantley L C, Neel B G, Lodish H F. Cell. 1995;80:729–738. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
