

The role of temporal cortical areas in perceptual organization
- PMID: 9096407
- PMCID: PMC20383
- DOI: 10.1073/pnas.94.7.3408
The role of temporal cortical areas in perceptual organization
Abstract
The visual areas of the temporal lobe of the primate are thought to be essential for the representation of visual objects. To examine the role of these areas in the visual awareness of a stimulus, we recorded the activity of single neurons in monkeys trained to report their percepts when viewing ambiguous stimuli. Visual ambiguity was induced by presenting incongruent images to the two eyes, a stimulation condition known to instigate binocular rivalry, during which one image is seen at a given time while the other is perceptually suppressed. Previous recordings in areas V1, V2, V4, and MT of monkeys experiencing binocular rivalry showed that only a small proportion of striate and early extrastriate neurons discharge exclusively when the driving stimulus is seen. In contrast, the activity of almost all neurons in the inferior temporal cortex and the visual areas of the cortex of superior temporal sulcus was found to be contingent upon the perceptual dominance of an effective visual stimulus. These areas thus appear to represent a stage of processing beyond the resolution of ambiguities--and thus beyond the processes of perceptual grouping and image segmentation--where neural activity reflects the brain's internal view of objects, rather than the effects of the retinal stimulus on cells encoding simple visual features or shape primitives.
Figures

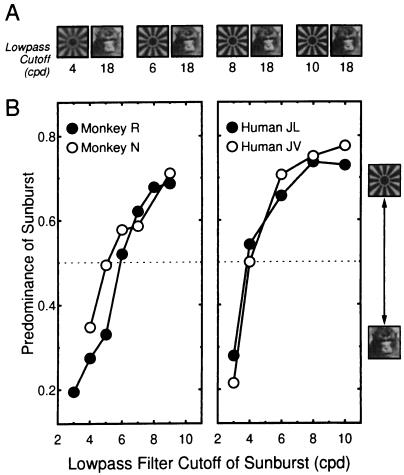
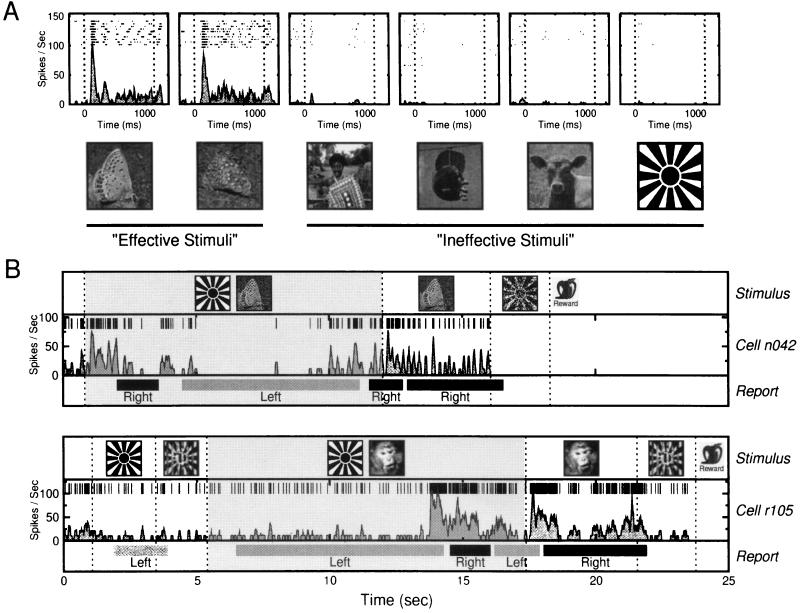
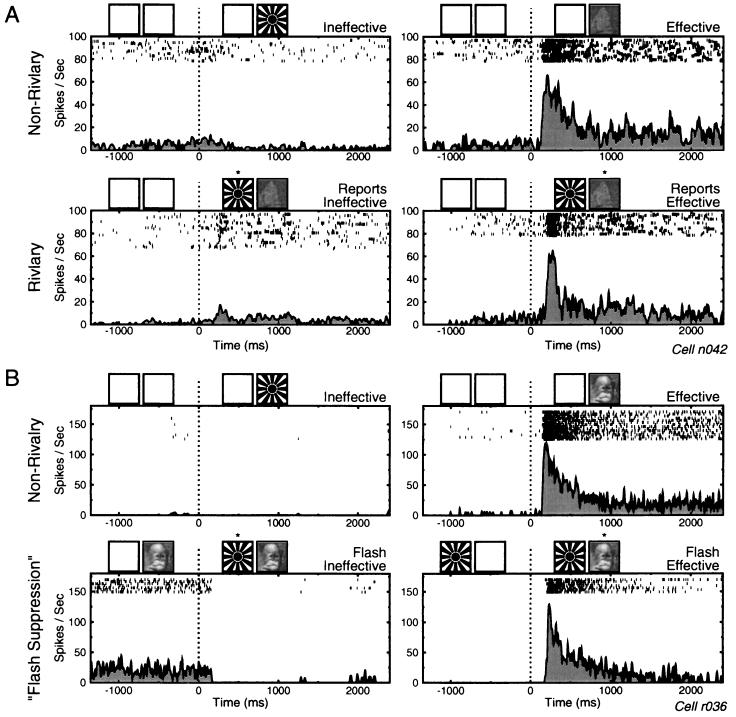
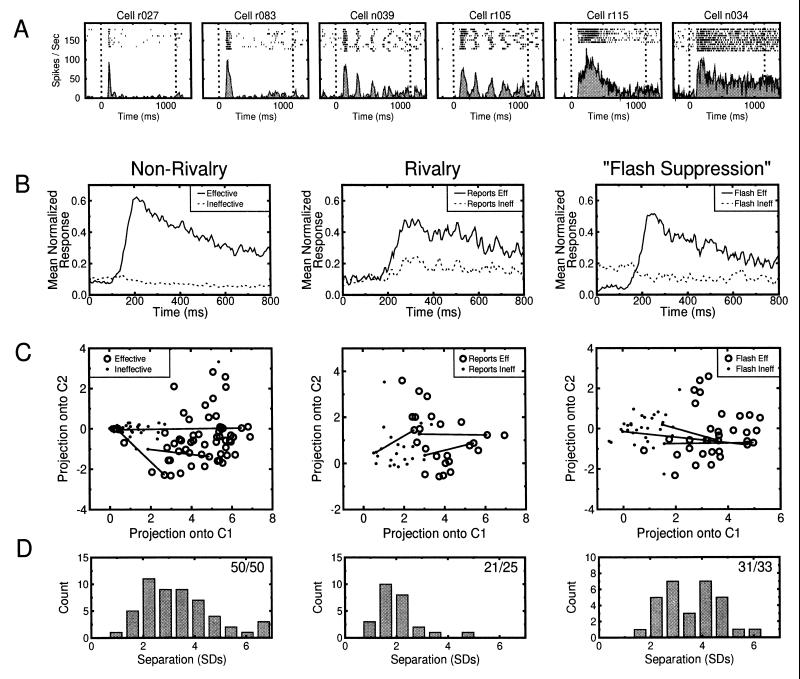
References
-
- Desimone R, Duncan J. Annu Rev Neurosci. 1995;18:193–222. - PubMed
-
- Logothetis N K, Sheinberg D L. Annu Rev Neurosci. 1996;19:577–621. - PubMed
-
- Logothetis N K, Leopold D A, Sheinberg D L. Nature (London) 1996;380:621–624. - PubMed
-
- Blake R R. Psychol Rev. 1989;96:145–167. - PubMed
-
- Leopold D A, Logothetis N K. Nature (London) 1996;379:549–553. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
