

Mammary-derived signals activate programmed cell death during the first stage of mammary gland involution
- PMID: 9096410
- PMCID: PMC20386
- DOI: 10.1073/pnas.94.7.3425
Mammary-derived signals activate programmed cell death during the first stage of mammary gland involution
Abstract
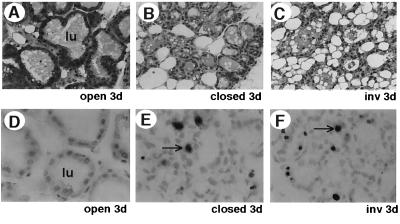
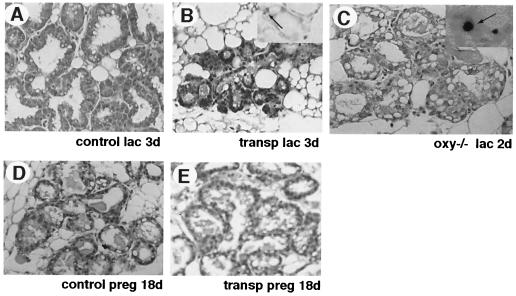
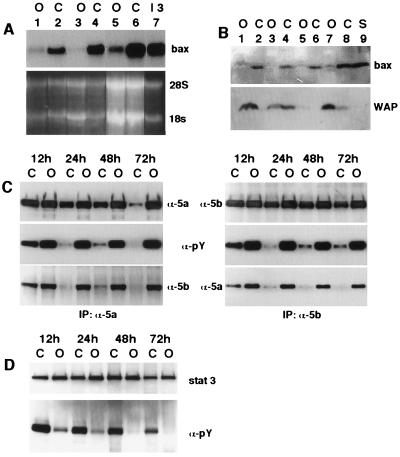
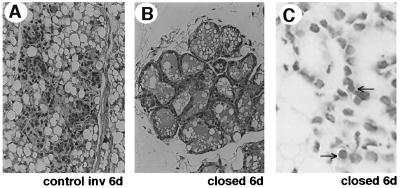
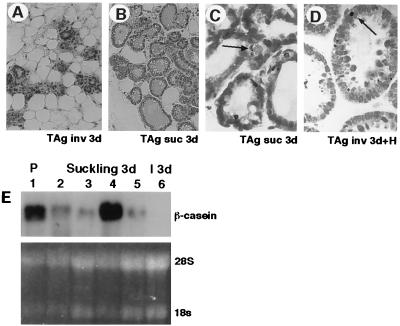
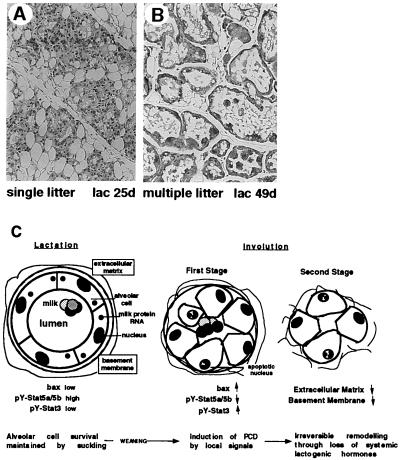
Programmed cell death (PCD) of mammary alveolar cells during involution commences within hours of the end of suckling. Locally, milk accumulates within alveolar lumens; systemically, levels of lactogenic hormones fall. Four experimental models were used to define the role of local factors as compared with systemic hormones during the first and second stages of involution. In three models, milk release was disrupted in the presence of systemic lactogenic hormones: (i) sealing of the teats, (ii) mammary gland transplants that cannot release milk due to the absence of a teat connection, and (iii) inactivation of the oxytocin gene. The ability of systemic hormones to preserve lobular-alveolar structure without blocking PCD was illustrated using a fourth transgenic model of lactation failure. During the first stage of involution, local signals were sufficient to induce alveolar PCD even in the presence of systemic lactogenic hormones. PCD coincided with bax induction, decreased expression of milk proteins, block of prolactin signal transduction through Stat5a and 5b, and activation of Stat3. The two stages of mammary gland involution are regulated by progressive gain of death signals and loss of survival factors. This study demonstrates that genetic events that occur during the first reversible stage are controlled by local factors. These mammary-derived death signals are dominant over protective effects related to systemic hormone stimulation.
Figures
References
-
- Strange R, Li F, Saurer S, Burkhardt A, Friis R. Development. 1992;115:49–58. - PubMed
-
- Marti A, Jehn B, Costello E, Keon N, Ke G, Martin F, Jaggi R. Oncogene. 1994;9:1213–1223. - PubMed
-
- Marti A, Feng Z, Jehn B, Djonov V, Chicaiza G, Altermatt H J, Jaggi R. Cell Death Differ. 1995;2:277–283. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
