

Lack of mitochondrial and nuclear-encoded subunits of complex I and alteration of the respiratory chain in Nicotiana sylvestris mitochondrial deletion mutants
- PMID: 9096412
- PMCID: PMC20388
- DOI: 10.1073/pnas.94.7.3436
Lack of mitochondrial and nuclear-encoded subunits of complex I and alteration of the respiratory chain in Nicotiana sylvestris mitochondrial deletion mutants
Abstract
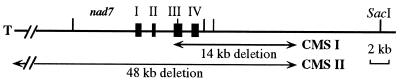
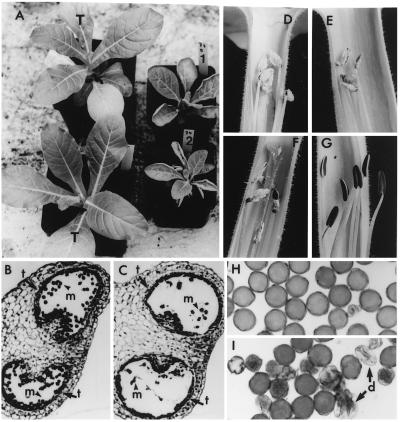
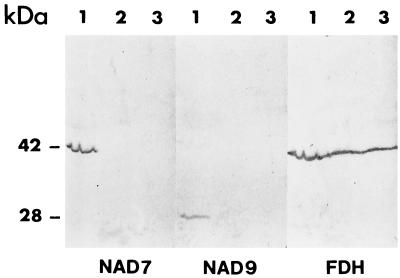
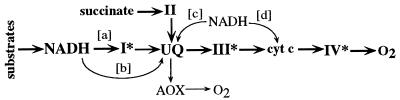
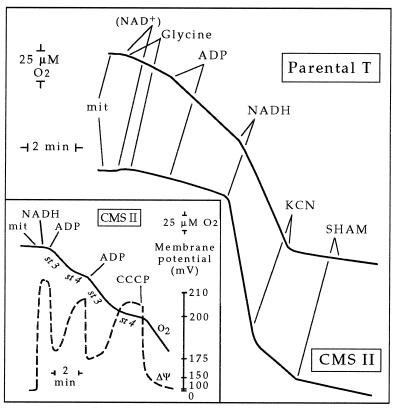
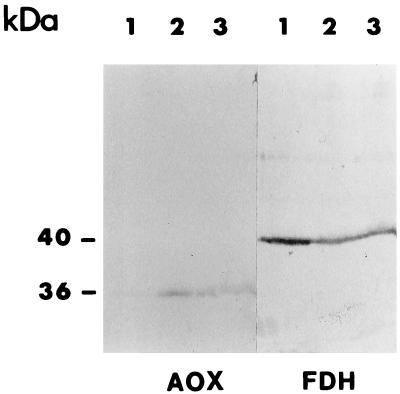
We previously have shown that Nicotiana sylvestris cytoplasmic male sterile (CMS) mutants I and II present large mtDNA deletions and that the NAD7 subunit of complex I (the main dehydrogenase of the mitochondrial respiratory chain) is absent in CMS I. Here, we show that, despite a large difference in size in the mtDNA deletion, CMS I and II display similar alterations. Both have an impaired development from germination to flowering, with partial male sterility that becomes complete under low light. Besides NAD7, two other complex I subunits are missing (NAD9 and the nucleus-encoded, 38-kDa subunit), identified on two-dimensional patterns of mitochondrial proteins. Mitochondria isolated from CMS leaves showed altered respiration. Although their succinate oxidation through complex II was close to that of the wild type, oxidation of glycine, a priority substrate of plant mitochondria, was significantly reduced. The remaining activity was much less sensitive to rotenone, indicating the breakdown of Complex I activity. Oxidation of exogenous NADH (coupled to proton gradient generation and partly sensitive to rotenone) was strongly increased. These results suggest respiratory compensation mechanisms involving additional NADH dehydrogenases to complex I. Finally, the capacity of the cyanide-resistant alternative oxidase pathway was enhanced in CMS, and higher amounts of enzyme were evidenced by immunodetection.
Figures


References
-
- Schuster W, Brennicke A. Annu Rev Plant Physiol Plant Mol Biol. 1994;45:61–78.
-
- Walker J E. Q Rev Biophys. 1992;25:253–324. - PubMed
-
- Weiss H, Friedrich T, Hofhaus G, Preis D. Eur J Biochem. 1991;197:563–576. - PubMed
-
- Chomyn A, Cleeter M W J, Ragan C I, Riley M, Doolitle R F, Attardi G. Science. 1986;234:614–618. - PubMed
-
- Lamattina L, Gonzalez D, Gualberto J, Grienenberger J M. Eur J Biochem. 1993;217:831–838. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
