

Monovalent cation permeation through the connexin40 gap junction channel. Cs, Rb, K, Na, Li, TEA, TMA, TBA, and effects of anions Br, Cl, F, acetate, aspartate, glutamate, and NO3
- PMID: 9101408
- PMCID: PMC2219432
- DOI: 10.1085/jgp.109.4.509
Monovalent cation permeation through the connexin40 gap junction channel. Cs, Rb, K, Na, Li, TEA, TMA, TBA, and effects of anions Br, Cl, F, acetate, aspartate, glutamate, and NO3
Abstract
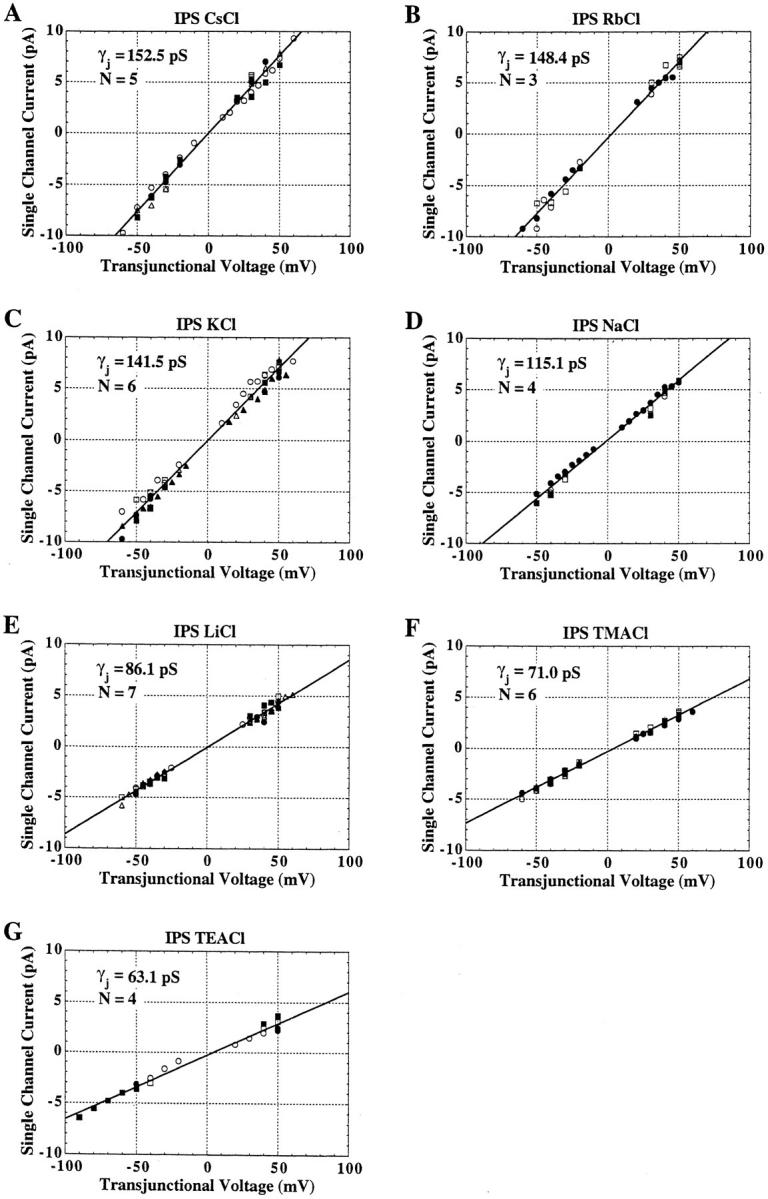
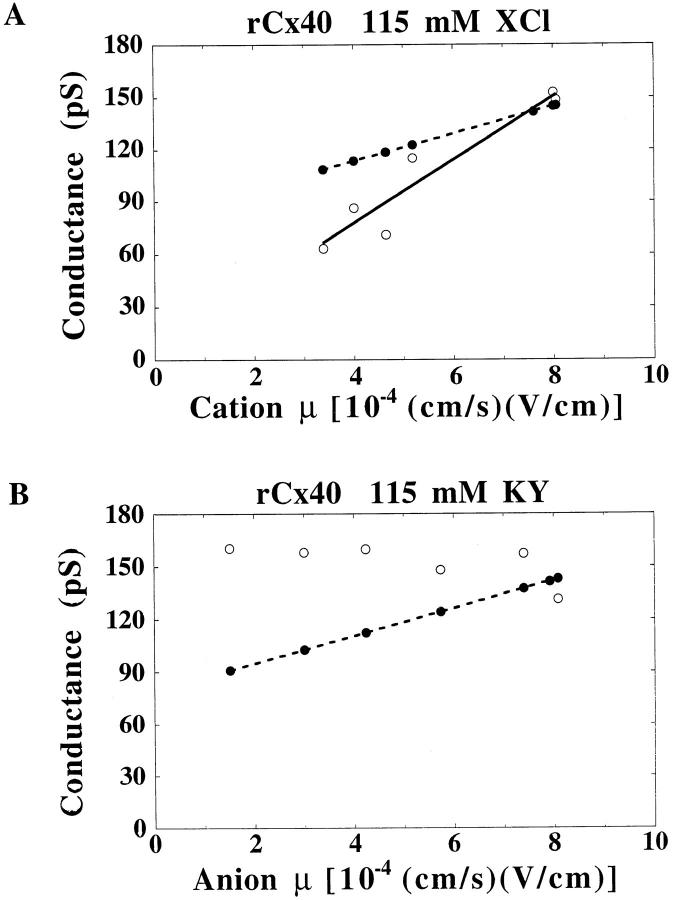
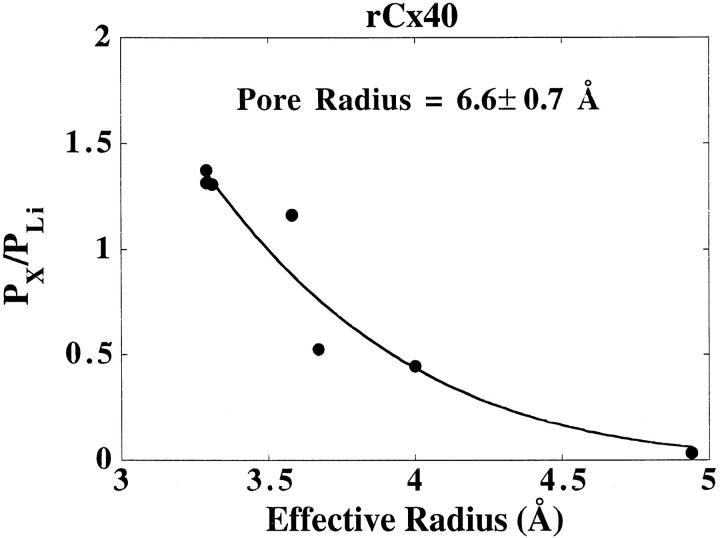
The unitary conductances and permeability sequences of the rat connexin40 (rCx40) gap junction channels to seven monovalent cations and anions were studied in rCx40-transfected neuroblastoma 2A (N2A) cell pairs using the dual whole cell recording technique. Chloride salt cation substitutions (115 mM principal salt) resulted in the following junctional maximal single channel current-voltage relationship slope conductances (gamma 1 in pS): CsC1 (153), RbC1 (148), KC1 (142), NaC1 (115), LiC1 (86), TMAC1 (71), TEAC1 (63). Reversible block of the rCx40 channel was observed with TBA. Potassium anion salt gamma j are: Kglutamate (160), Kacetate (160), Kaspartate (158), KNO3 (157), KF (148), KC1 (142), and KBr (132). Ion selectivity was verified by measuring reversal potentials for current in rCx40 gap junction channels with asymmetric salt solutions in the two electrodes and using the Goldman-Hodgkin-Katz equation to calculate relative permeabilities. The permeabilities relative to Li+ are: Cs+ (1.38), Rb+ (1.32), K+ (1.31), Na+ (1.16), TMA+ (0.53), TEA+ (0.45), TBA+ (0.03), Cl- (0.19), glutamate+ (0.04), and NO(3)- (0.14), assuming that the monovalent anions permeate the channel by forming ion pairs with permeant monovalent cations within the pore thereby causing proportionate decreases in the channel conductance. This hypothesis can account for why the predicted increasing conductances with increasing ion mobilities in an essentially aqueous channel were not observed for anions in the rCx40 channel. The rCx40 effective channel radius is estimated to be 6.6 A from a theoretical fit of the relationship of relative permeability and cation radius.
Figures




References
-
- Andersen, O.S. 1975. Ion-specificity of gramicidin channels. In International Biophysics Congress. Copenhagen, Denmark. 112. (Abstr.).
-
- Beblo DA, Wang H-Z, Beyer EC, Westphale EM, Veenstra RD. Unique conductance, gating, and selective permeability properties of gap junction channels formed by connexin40. Circ Res. 1995;77:813–822. - PubMed
-
- Bennett, M.V.L., and D.C. Spray. 1965. Gap Junctions. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
