

Bystander activation of cytotoxic T cells: studies on the mechanism and evaluation of in vivo significance in a transgenic mouse model
- PMID: 9104811
- PMCID: PMC2196250
- DOI: 10.1084/jem.185.7.1241
Bystander activation of cytotoxic T cells: studies on the mechanism and evaluation of in vivo significance in a transgenic mouse model
Abstract
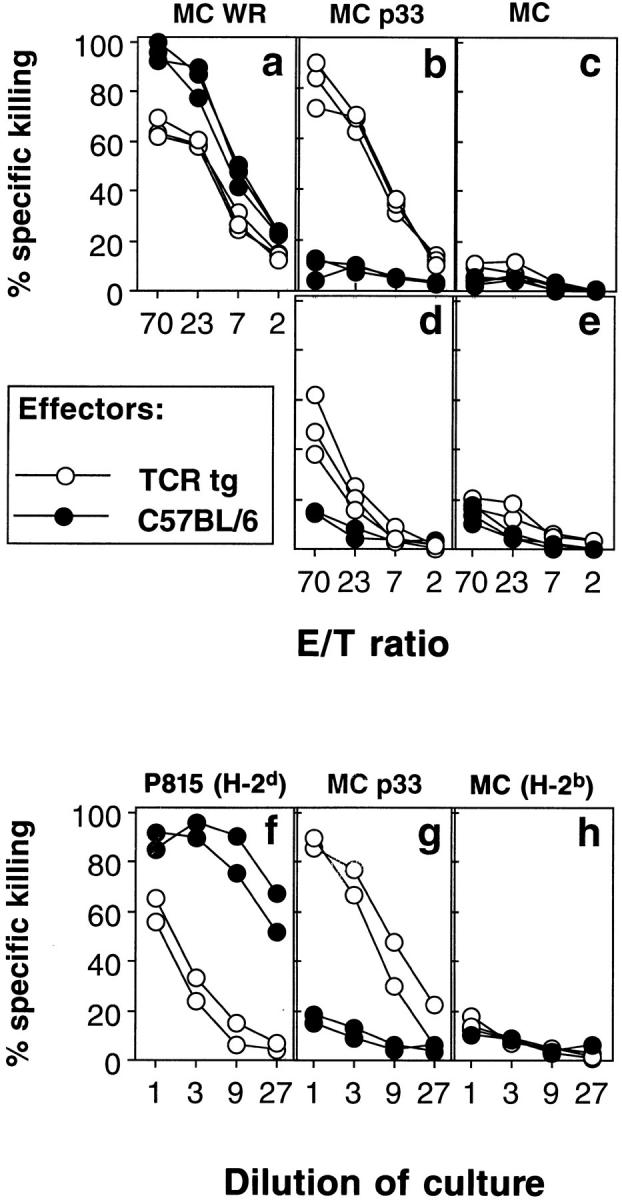
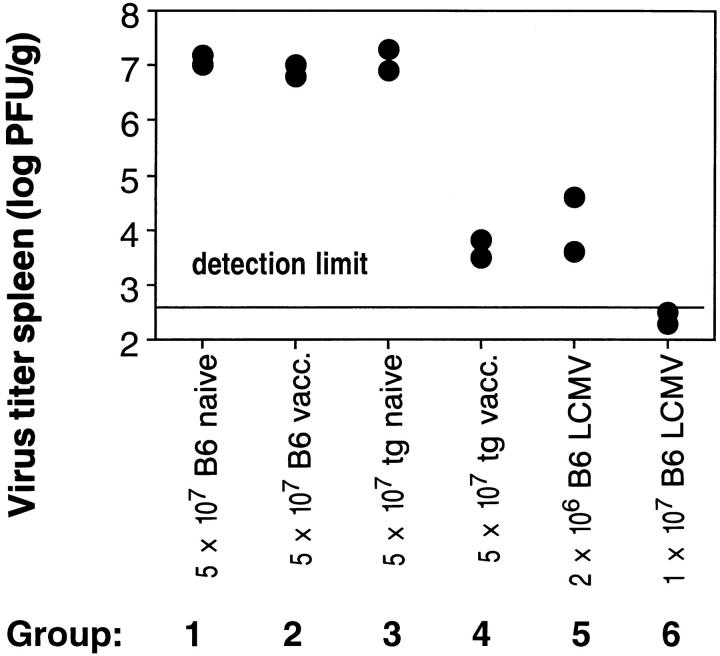
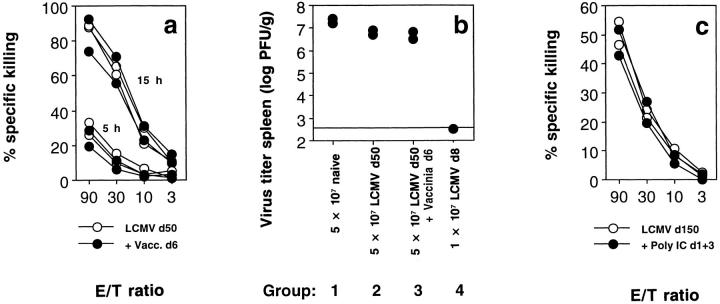
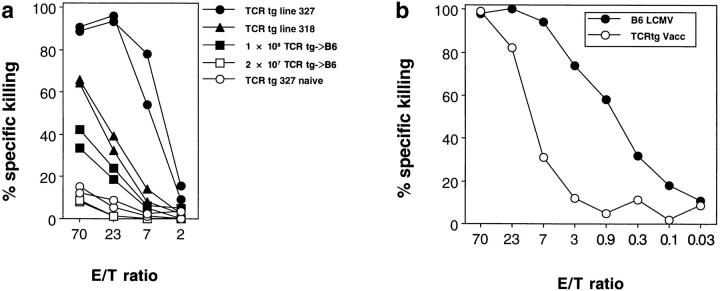
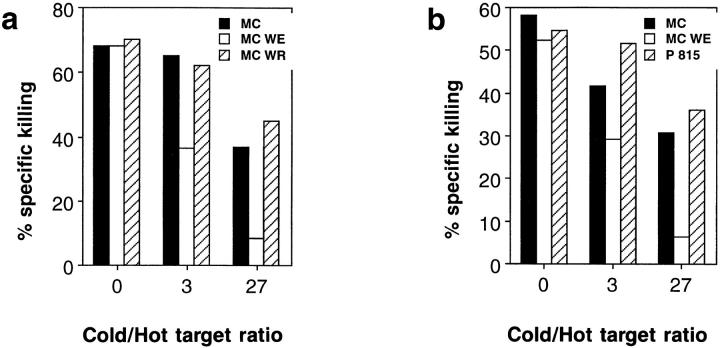
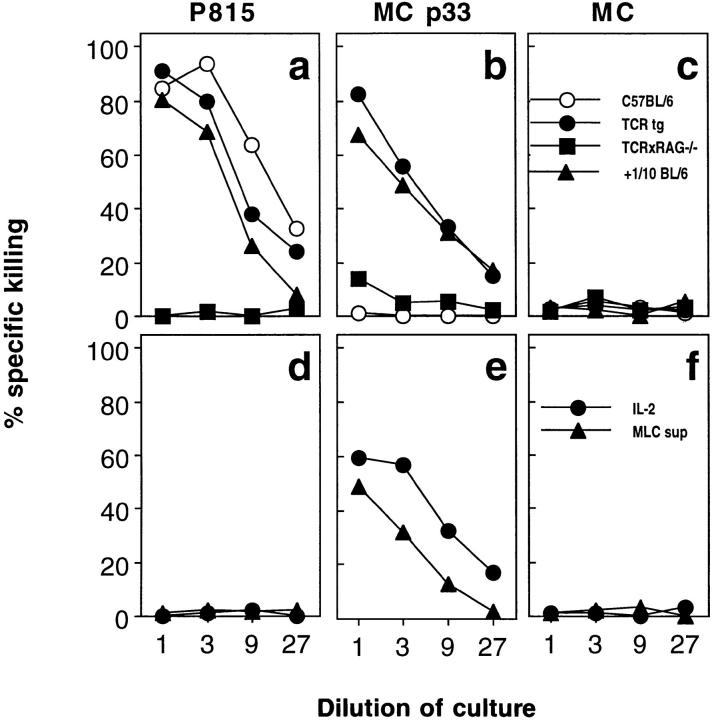
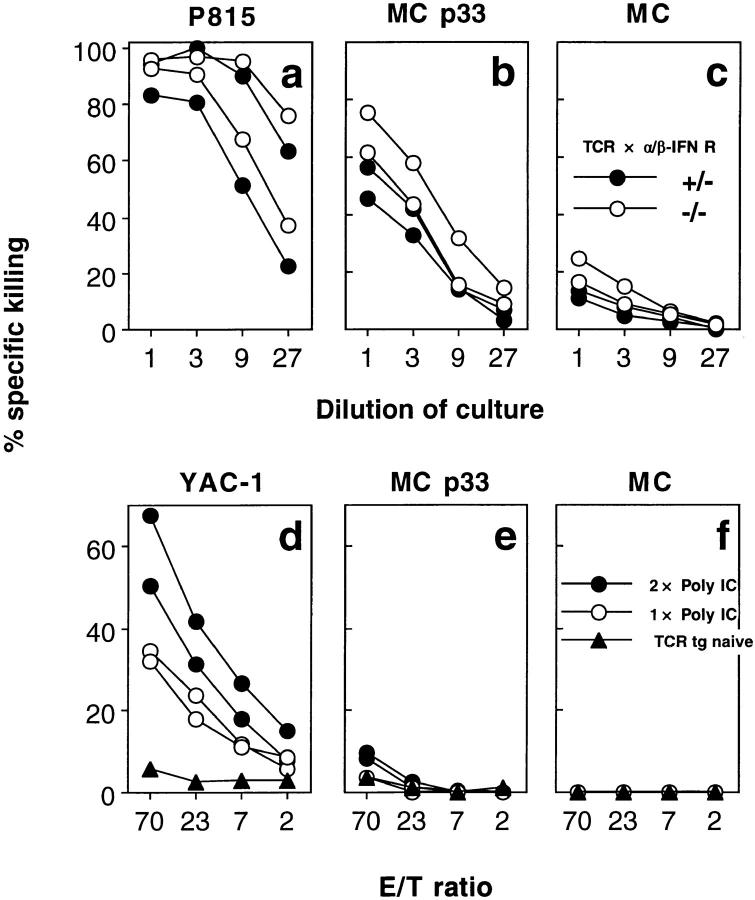
Bystander activation, i.e., activation of T cells specific for an antigen X during an immune response against antigen Y may occur during viral infections. However, the low frequency of bystander-activated T cells has rendered it difficult to define the mechanisms and possible in vivo relevance of this nonspecific activation. This study uses transgenic mice expressing a major histocompatibility complex class I-restricted TCR specific for glycoprotein peptide 33-41 of lymphocytic choriomeningitis virus (LCMV) to overcome this limitation. CD8+ T cells from specific pathogen-free maintained, unimmunized "naive" TCR transgenic mice can differentiate into LCMV-specific cytolytic effector CTL during infections with vaccinia virus or Listeria monocytogenes in vivo or mixed lymphocyte culture in vitro. We show that in these model situations (a) nonspecifically activated CTL are able to confer antiviral protection in vivo, (b) bystander activation is largely independent of the expression of a second T cell receptor of different specificity, (c) bystander activation is not mediated by a broadly cross-reactive TCR, but rather by cytokines, (d) bystander activation can be mediated by cytokines such as IL-2, but not alpha/beta-IFN in vitro; (e) bystander activation is, overall, a rare event, occuring in vivo in roughly 1 in 200 of the LCMV-specific CTL during infection of TCR transgenic mice with vaccinia virus; (f) bystander activation does not have a significant functional impact on nontransgenic CTL memory under the conditions tested; and (g) even in the TCR transgenic situation, where unphysiologically high numbers of T cells of a single specificity are present, bystander activation is not sufficient to cause clinically manifest autoimmune disease in a transgenic mouse model of diabetes. We conclude that although bystander activation via cytokines may generate cytolytically active CTL from naive precursors, quantitative considerations suggest that this is usually not of major biological consequence.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
