

High mobility group 1 protein is not stably associated with the chromosomes of somatic cells
- PMID: 9105033
- PMCID: PMC2139855
- DOI: 10.1083/jcb.137.1.19
High mobility group 1 protein is not stably associated with the chromosomes of somatic cells
Abstract
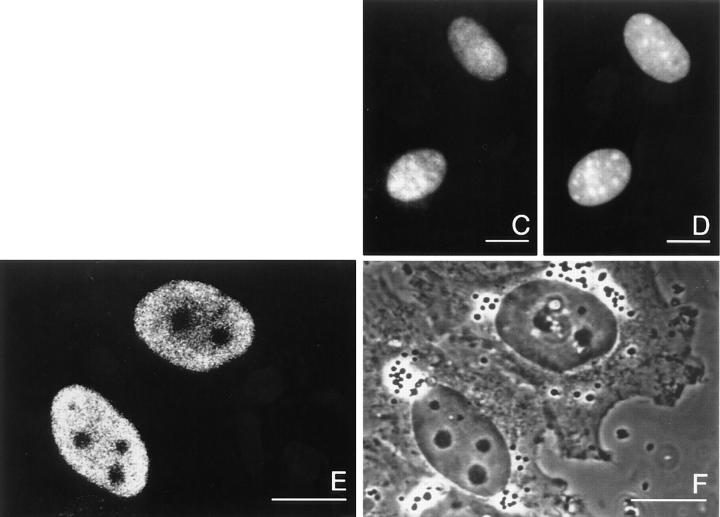
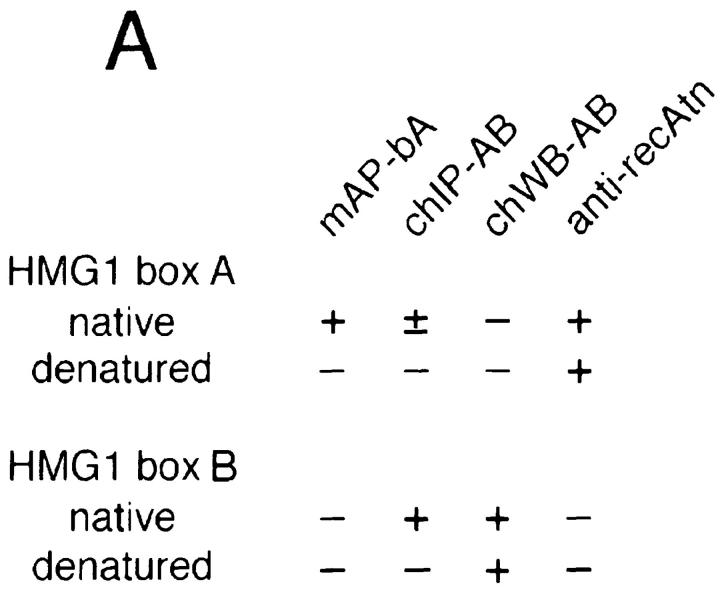
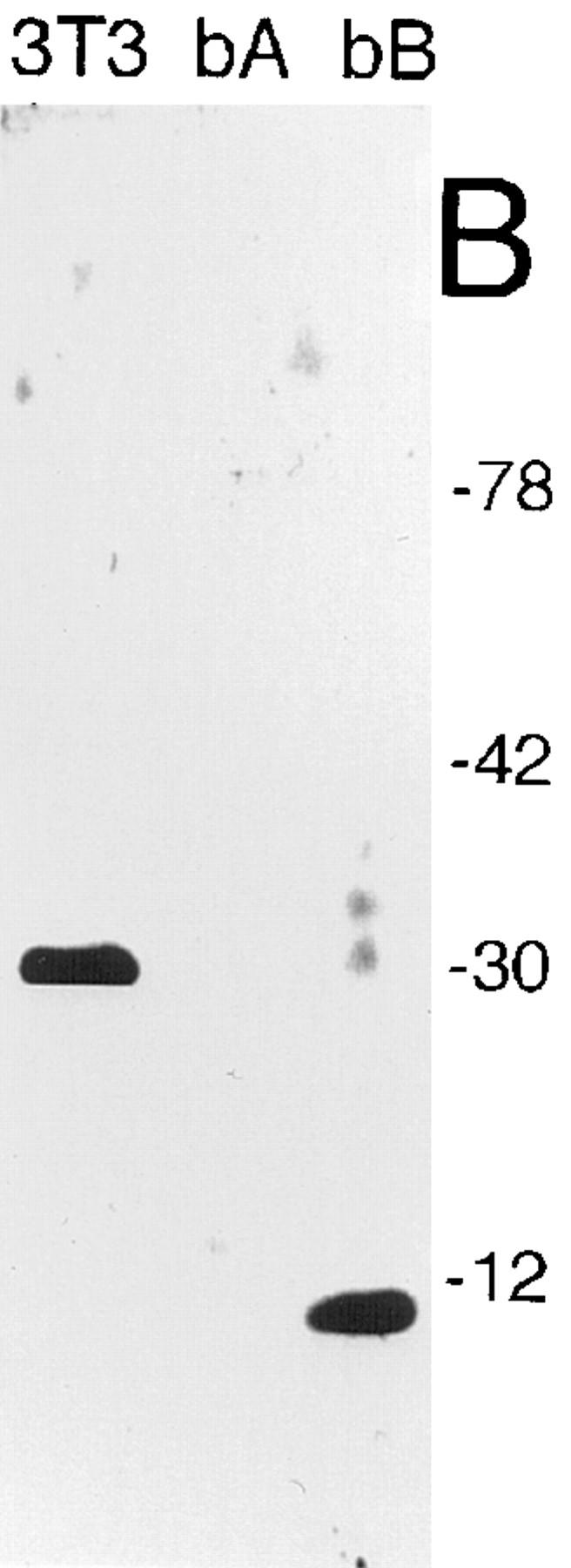
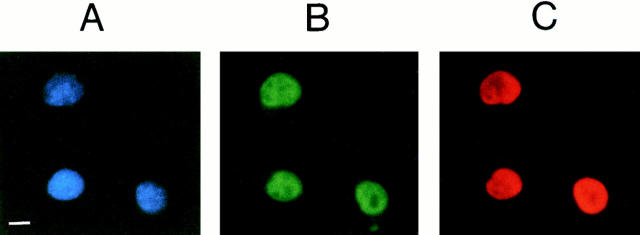
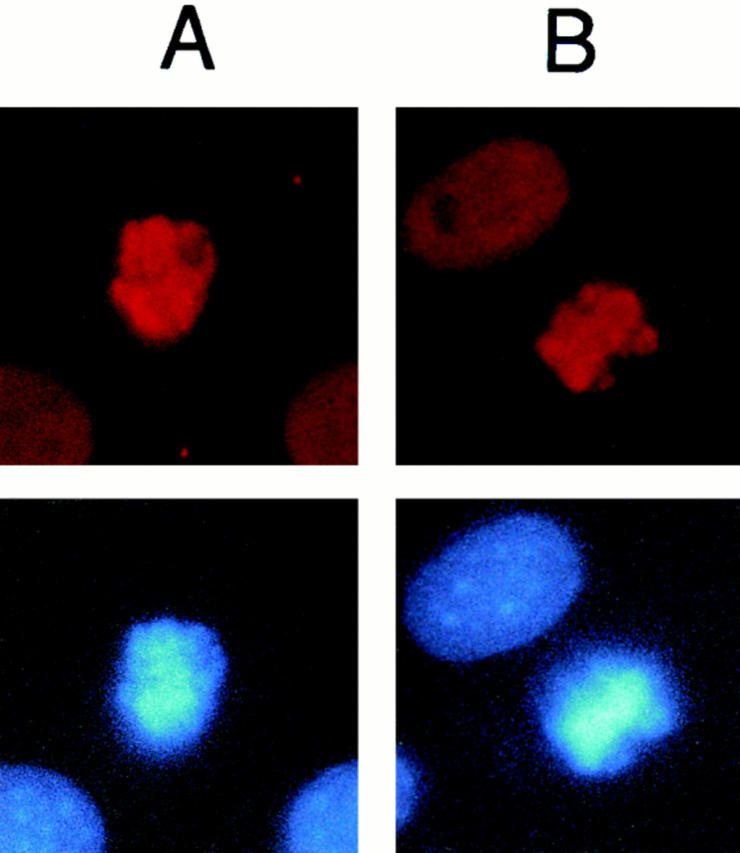
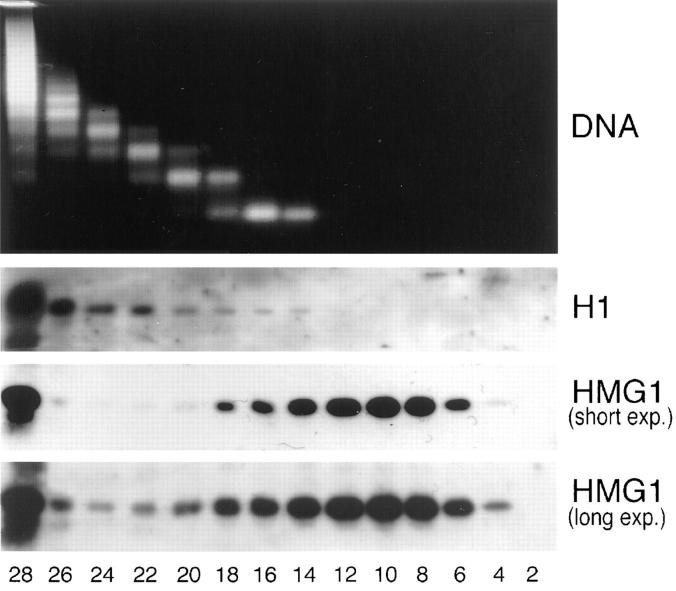
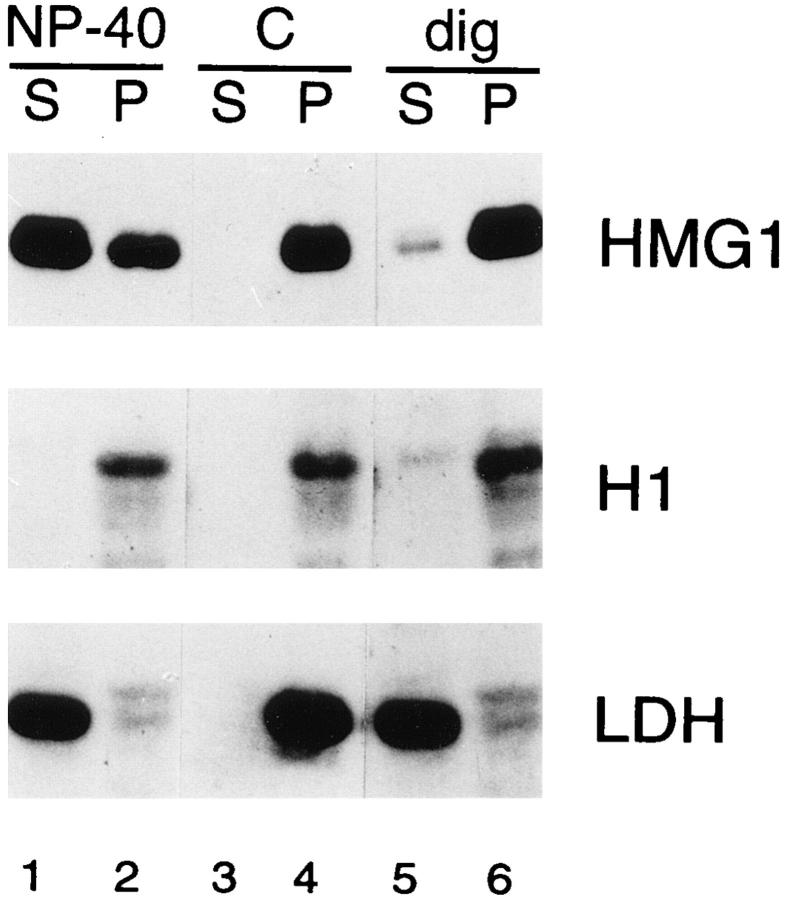
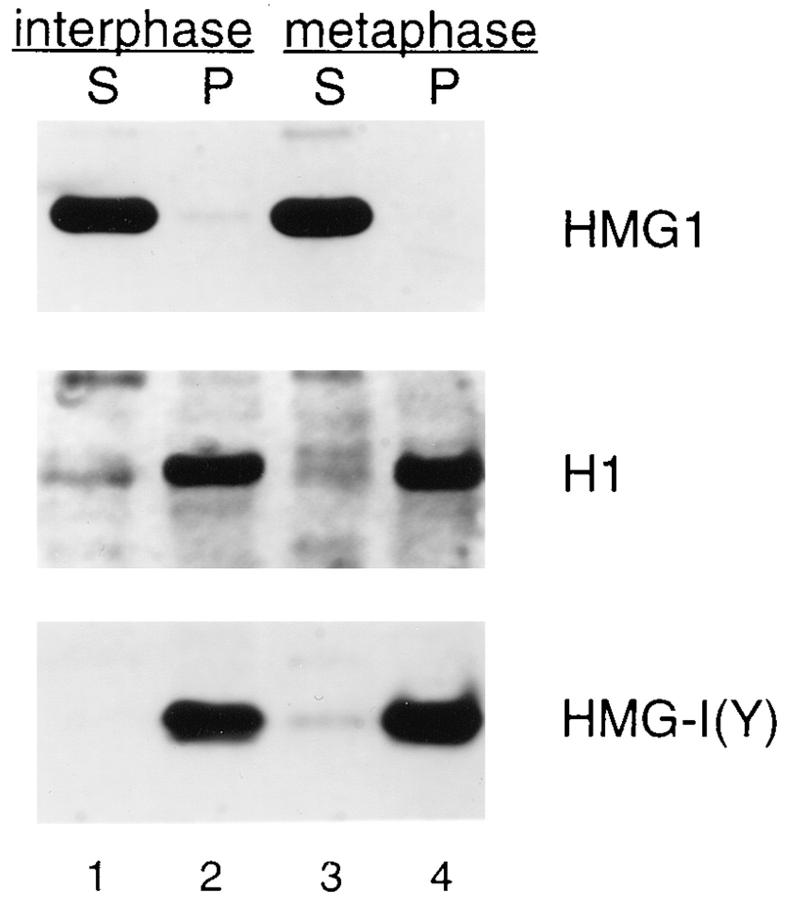
High mobility group 1 (HMG1) protein is an abundant and conserved component of vertebrate nuclei and has been proposed to play a structural role in chromatin organization, possibly similar to that of histone H1. However, a high abundance of HMG1 had also been reported in the cytoplasm and on the surface of mammalian cells. We conclusively show that HMG1 is a nuclear protein, since several different anti-HMG1 antibodies stain the nucleoplasm of cultured cells, and epitope-tagged HMG1 is localized in the nucleus only. The protein is excluded from nucleoli and is not associated to specific nuclear structures but rather appears to be uniformly distributed. HMG1 can bind in vitro to reconstituted core nucleosomes but is not stably associated to chromatin in live cells. At metaphase, HMG1 is detached from condensed chromosomes, contrary to histone H1. During interphase, HMG1 readily diffuses out of nuclei after permeabilization of the nuclear membranes with detergents, whereas histone H1 remains associated to chromatin. These properties exclude a shared function for HMG1 and H1 in differentiated cells, in spite of their similar biochemical properties. HMG1 may be stably associated only to a very minor population of nucleosomes or may interact transiently with nucleosomes during dynamic processes of chromatin remodeling.
Figures
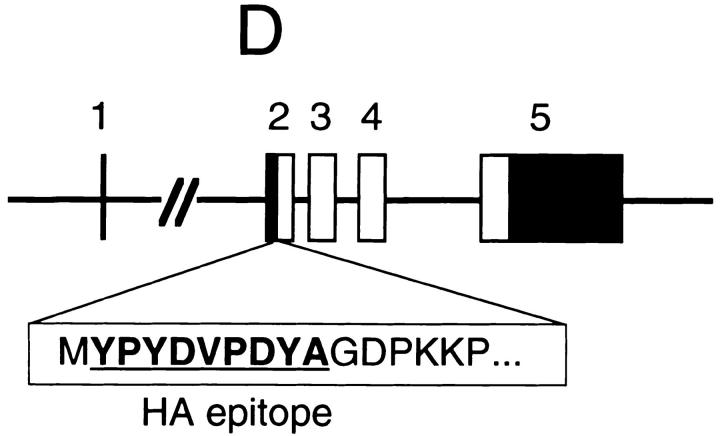
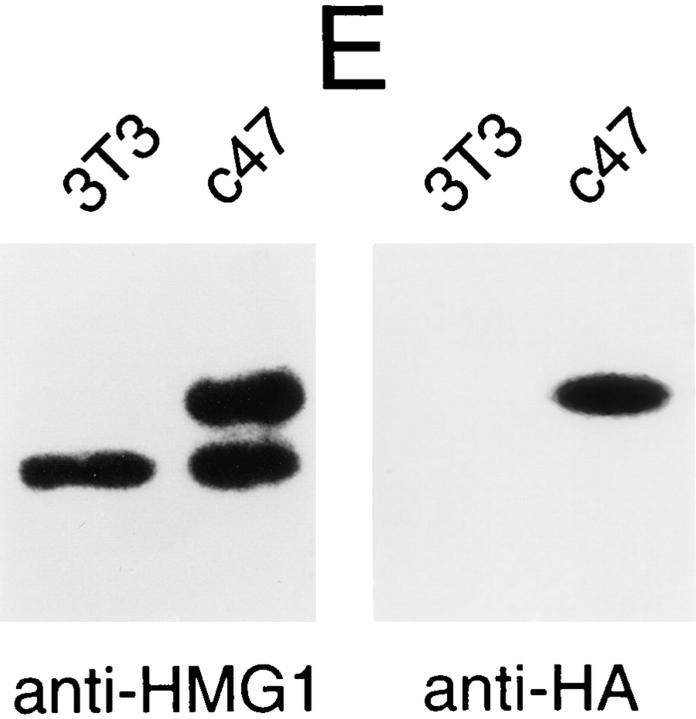
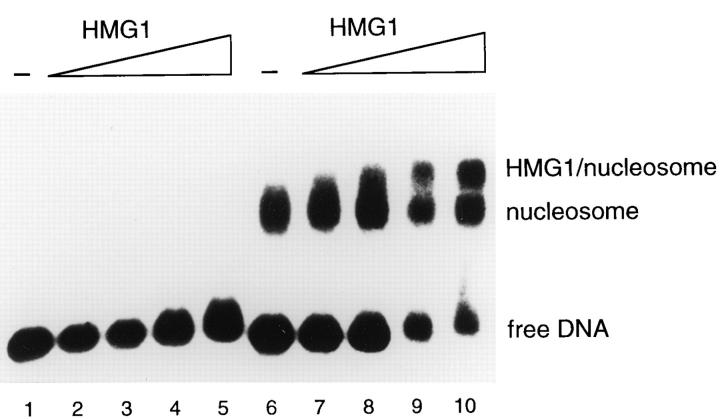
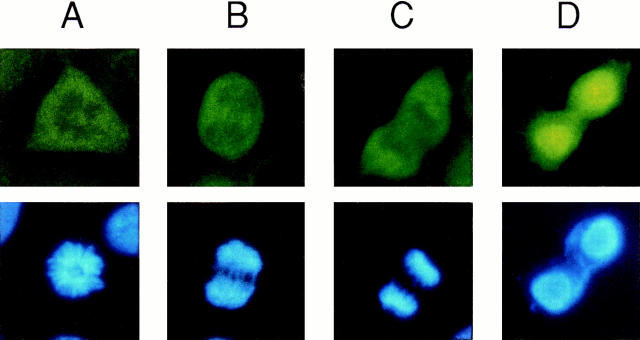
References
-
- Bianchi ME. Prokaryotic HU and eukaryotic HMG1: a kinked relationship. Mol Microbiol. 1994;14:1–5. - PubMed
-
- Bianchi, M.E. 1995. The HMG-box domain. In DNA–Protein: Structural Interactions. D.M.J. Lilley, editor. Oxford University Press, Oxford. 177–200.
-
- Bianchi ME, Beltrame M, Paonessa G. Specific recognition of cruciform DNA by nuclear protein HMG1. Science (Wash DC) 1989;243:1056–1059. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
