

The yeast gene, MDM20, is necessary for mitochondrial inheritance and organization of the actin cytoskeleton
- PMID: 9105043
- PMCID: PMC2139847
- DOI: 10.1083/jcb.137.1.141
The yeast gene, MDM20, is necessary for mitochondrial inheritance and organization of the actin cytoskeleton
Abstract
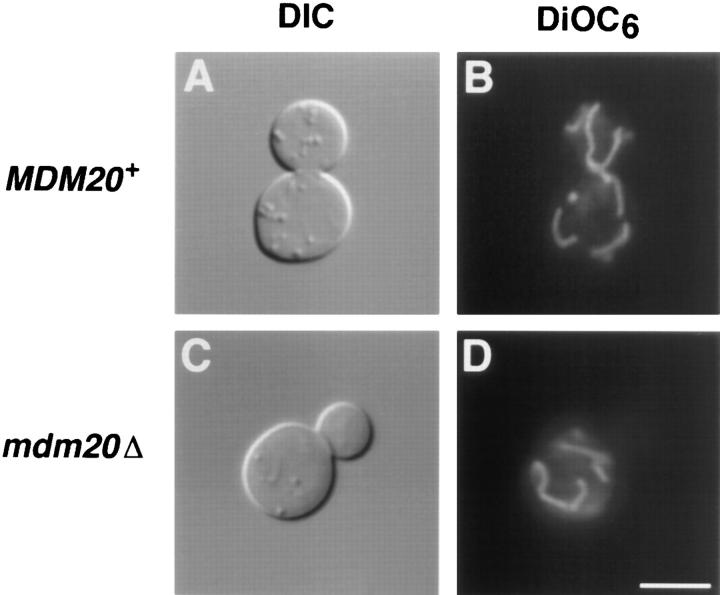
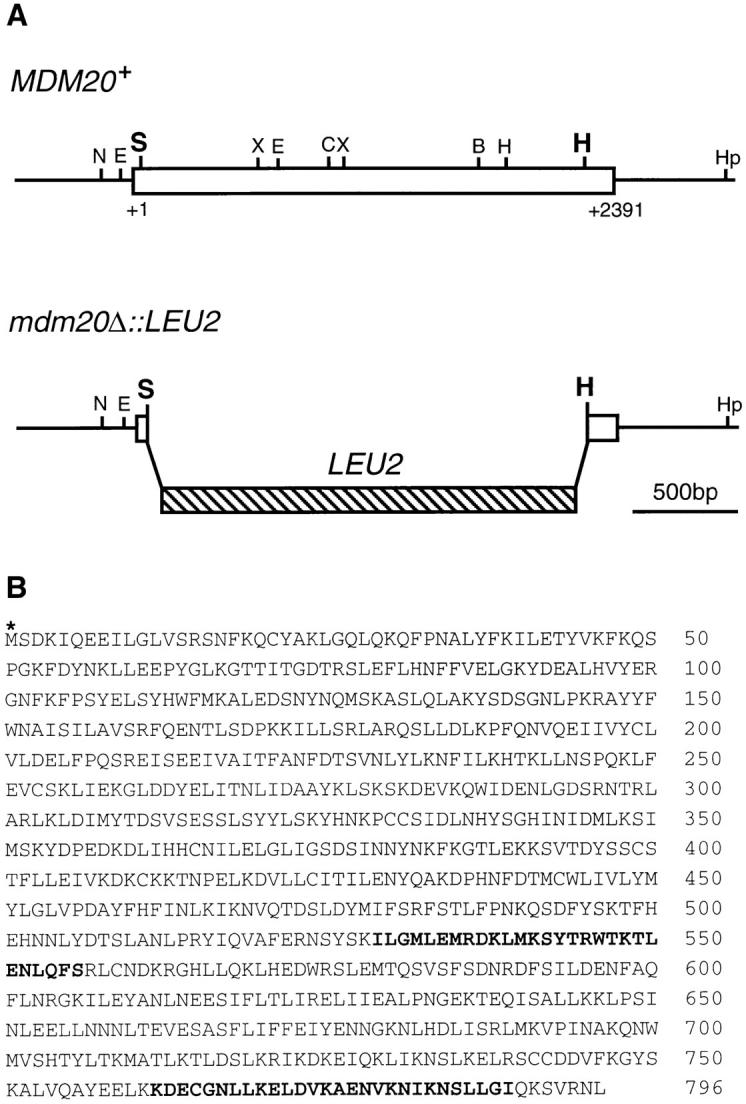
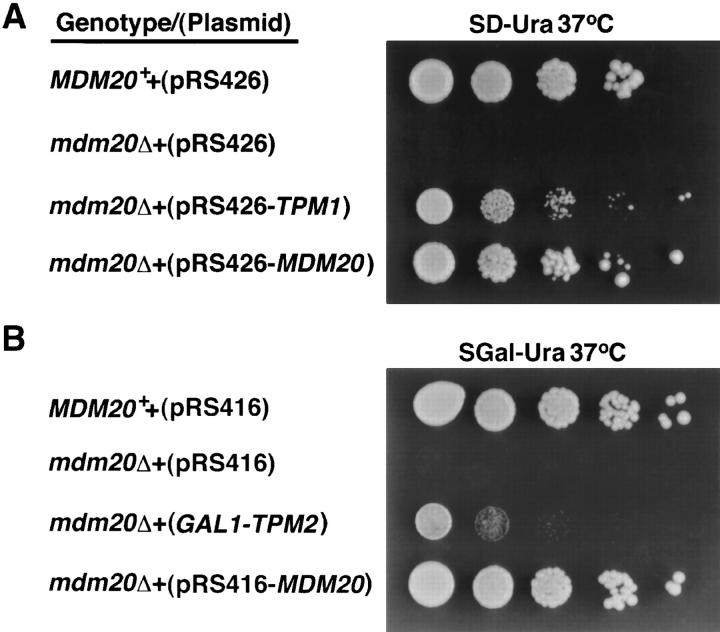
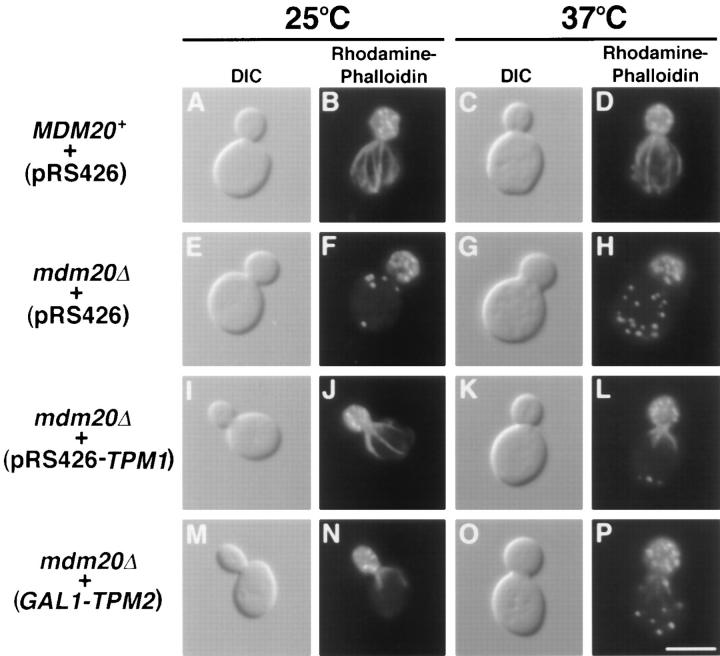
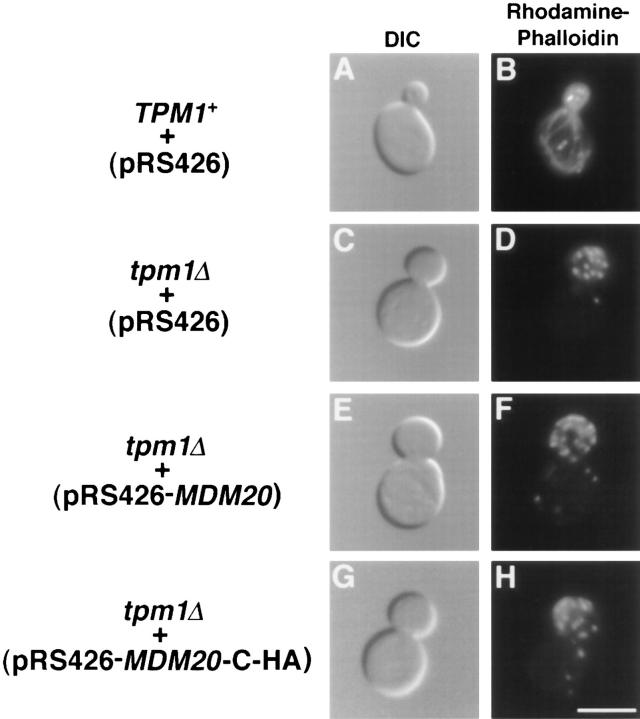
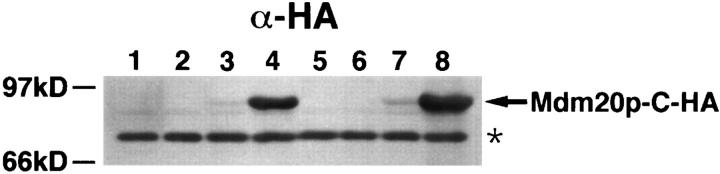
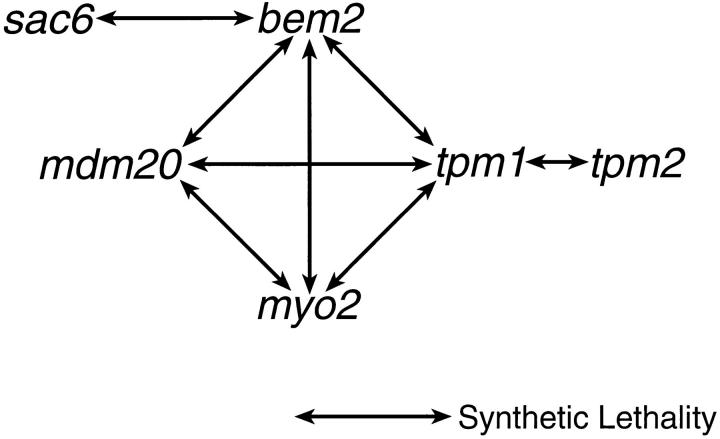
In Saccharomyces cerevisiae, the growing bud inherits a portion of the mitochondrial network from the mother cell soon after it emerges. Although this polarized transport of mitochondria is thought to require functions of the cytoskeleton, there are conflicting reports concerning the nature of the cytoskeletal element involved. Here we report the isolation of a yeast mutant, mdm20, in which both mitochondrial inheritance and actin cables (bundles of actin filaments) are disrupted. The MDM20 gene encodes a 93-kD polypeptide with no homology to other characterized proteins. Extra copies of TPM1, a gene encoding the actin filament-binding protein tropomyosin, suppress mitochondrial inheritance defects and partially restore actin cables in mdm20 delta cells. Synthetic lethality is also observed between mdm20 and tpm1 mutant strains. Overexpression of a second yeast tropomyosin, Tpm2p, rescues mutant phenotypes in the mdm20 strain to a lesser extent. Together, these results provide compelling evidence that mitochondrial inheritance in yeast is an actin-mediated process. MDM20 and TPM1 also exhibit the same pattern of genetic interactions; mutations in MDM20 are synthetically lethal with mutations in BEM2 and MYO2 but not SAC6. Although MDM20 and TPM1 are both required for the formation and/or stabilization of actin cables, mutations in these genes disrupt mitochondrial inheritance and nuclear segregation to different extents. Thus, Mdm20p and Tpm1p may act in vivo to establish molecular and functional heterogeneity of the actin cytoskeleton.
Figures
References
-
- Adams AEM, Pringle JR. Staining of actin with fluorochromeconjugated phalloidin. Methods Enzymol. 1991;194:729–731. - PubMed
-
- Adams AEM, Botstein D, Drubin DG. Requirement of yeast fimbrin for actin organization and morphogenesis in vivo. . Nature (Lond) 1991;354:404–408. - PubMed
-
- Altschul SF, Gish W, Miller W, Meyers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
