

Interleukin 1 beta-converting enzyme related proteases/caspases are involved in TRAIL-induced apoptosis of myeloma and leukemia cells
- PMID: 9105050
- PMCID: PMC2139852
- DOI: 10.1083/jcb.137.1.221
Interleukin 1 beta-converting enzyme related proteases/caspases are involved in TRAIL-induced apoptosis of myeloma and leukemia cells
Abstract
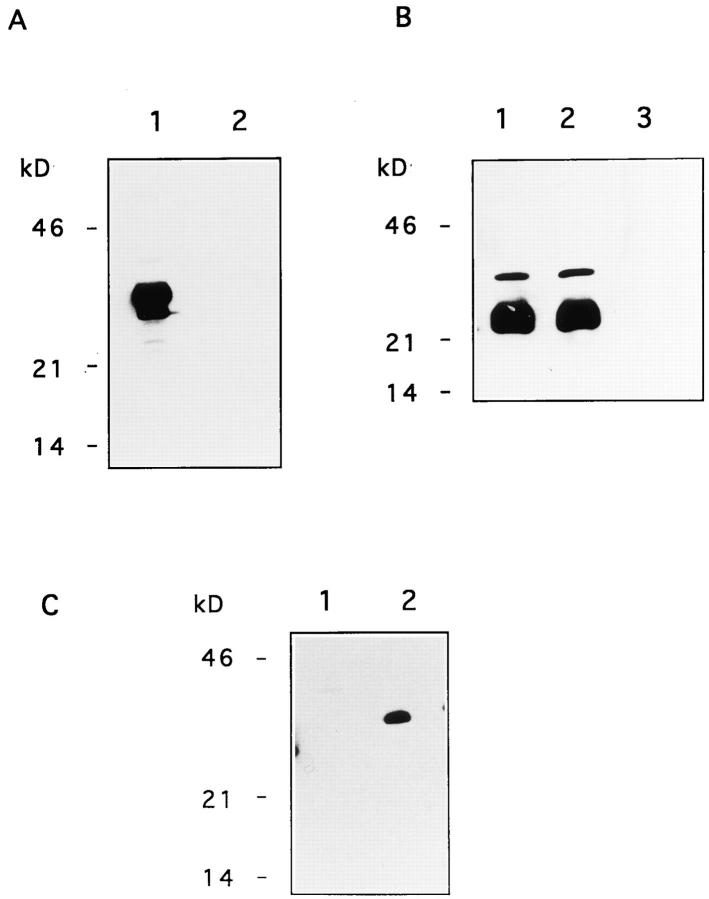
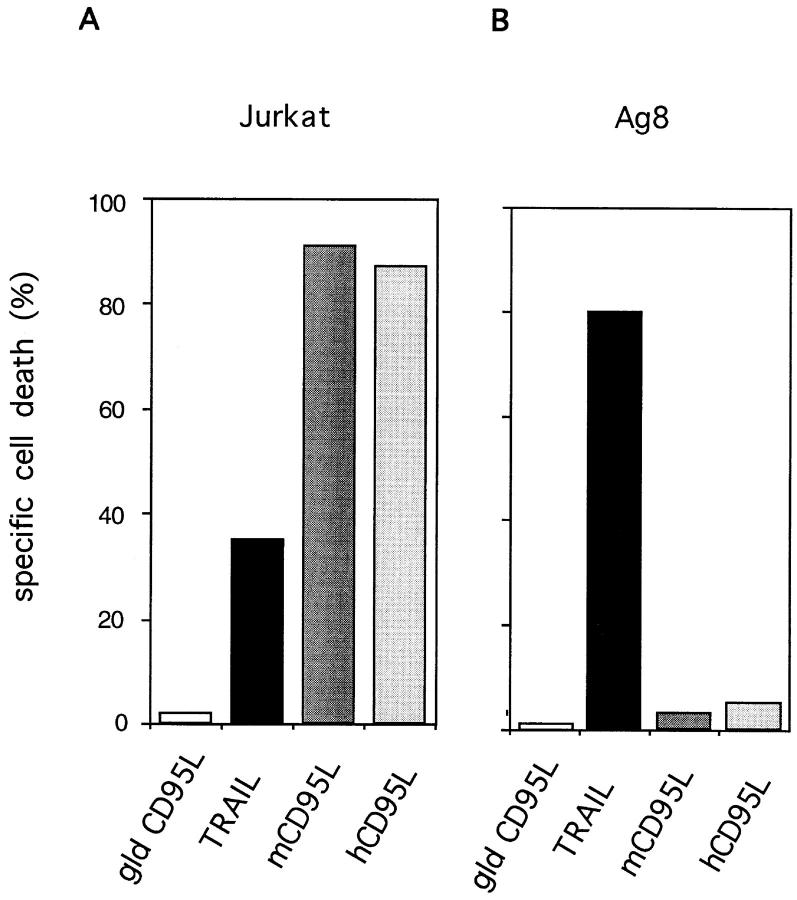
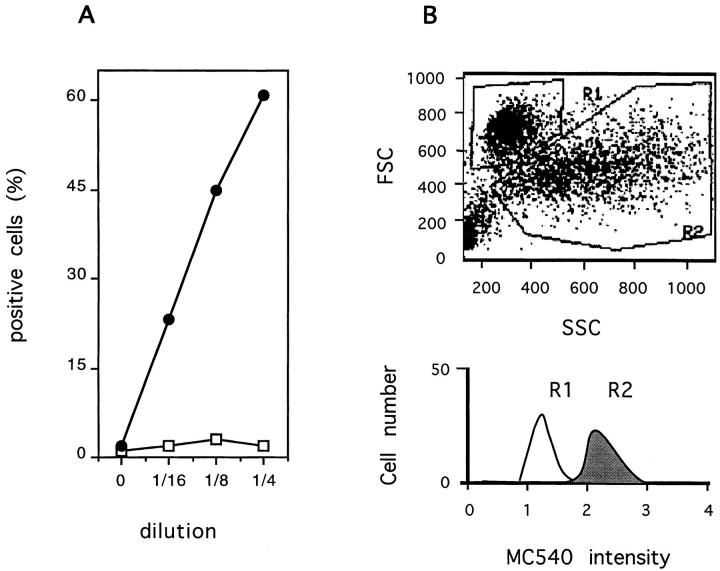
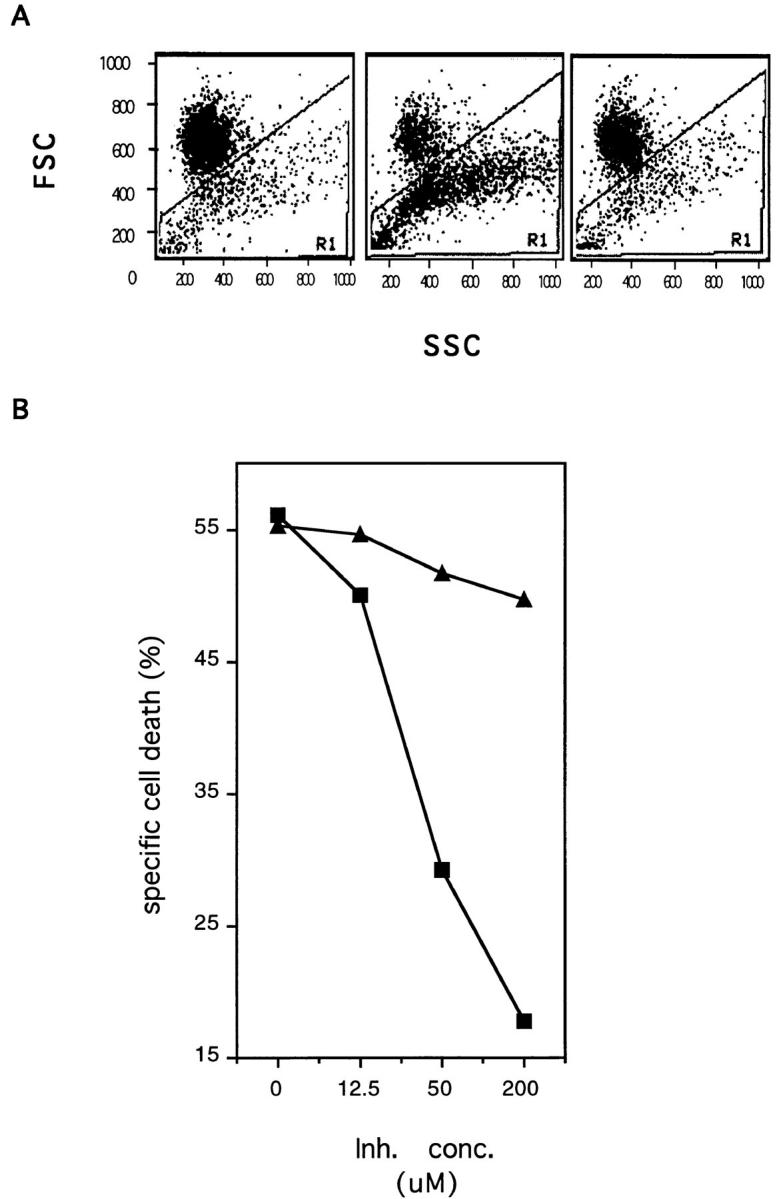
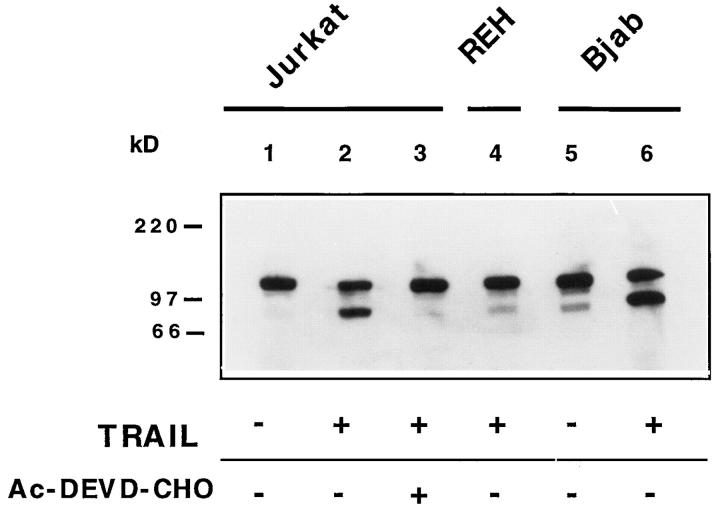
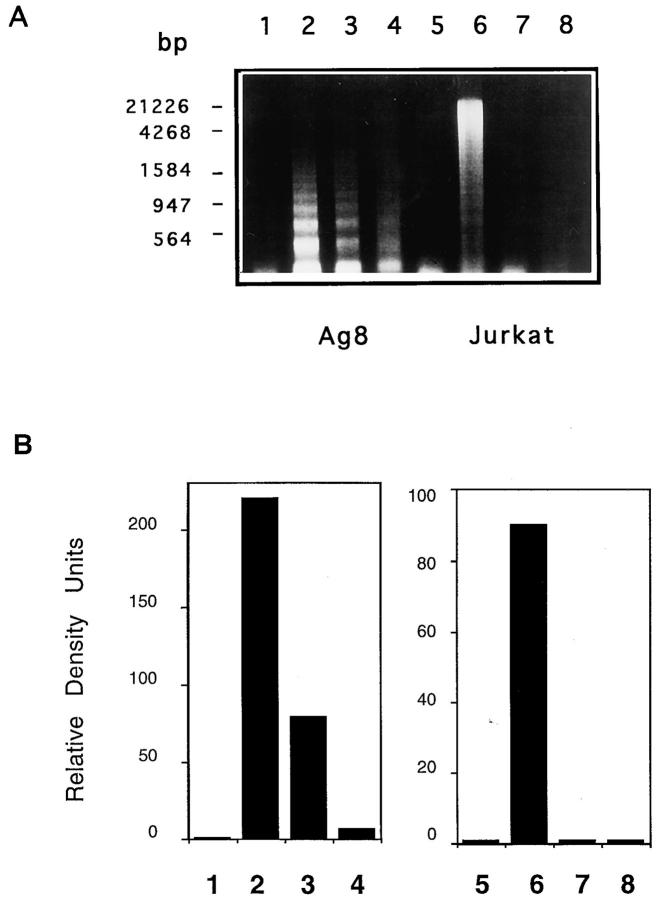
The Fas/APO-1/CD95 ligand (CD95L) and the recently cloned TRAIL ligand belong to the TNF-family and share the ability to induce apoptosis in sensitive target cells. Little information is available on the degree of functional redundancy between these two ligands in terms of target selectivity and intracellular signalling pathway(s). To address these issues, we have expressed and characterized recombinant mouse TRAIL. Specific detection with newly developed rabbit anti-TRAIL antibodies showed that the functional TRAIL molecule released into the supernatant of recombinant baculovirus-infected Sf9 cells is very similar to that associated with the membrane fraction of Sf9 cells. CD95L resistant myeloma cells were found to be sensitive to TRAIL, displaying apoptotic features similar to those of the CD95L- and TRAIL-sensitive T leukemia cells Jurkat. To assess if IL-1beta-converting enzyme (ICE) and/or ICE-related proteases (IRPs) (caspases) are involved in TRAIL-induced apoptosis of both cell types, peptide inhibition experiments were performed. The irreversible IRP/caspase-inhibitor Ac-YVAD-cmk and the reversible IRP/caspase-inhibitor Ac-DEVD-CHO blocked the morphological changes, disorganization of plasma membrane phospholipids, DNA fragmentation, and loss of cell viability associated with TRAIL-induced apoptosis. In addition, cells undergoing TRAIL-mediated apoptosis displayed cleavage of poly(ADP)-ribose polymerase (PARP) that was completely blocked by Ac-DEVD-CHO. These results indicate that TRAIL seems to complement the activity of the CD95 system as it allows cells, otherwise resistant, to undergo apoptosis triggered by specific extracellular ligands. Conversely, however, induction of apoptosis in sensitive cells by TRAIL involves IRPs/caspases in a fashion similar to CD95L. Thus, differential sensitivity to CD95L and TRAIL seems to map to the proximal signaling events associated with receptor triggering.
Figures



References
-
- Alnemri ES, Livingston DJ, Nicholson DW, Salvesen G, Thornberry NA, Wong WW, Yuan J. Human ICE/CED-3 protease nomenclature. Cell. 1996;87:171. - PubMed
-
- Boldin MP, Varfolomeev EE, Pancer Z, Mett IL, Camonis JH, Wallach D. A novel protein that interacts with the death domain of FAS/ APO-1 contains a sequence motif related to the death domain. J Biol Chem. 1995;270:7795–7798. - PubMed
-
- Boldin MP, Goncharov TM, Goltsev YV, Wallach D. Involvement of MACH, a novel MORT1/FADD-interacting protease, in Fas/APO-1- and TNF receptor-induced cell death. Cell. 1996;85:803–815. - PubMed
-
- Brunner T, Mogil RJ, LaFace D, Yoo NJ, Mahboubi A, Echeverry F, Martin SJ, Force WR, Lynch DH, Ware CF, et al. Cell-autonomous Fas (CD95)/Fas-ligand interaction mediates activation-induced apoptosis in T-cell hybridoma. Nature (Lond) 1995;373:441–444. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
