

Nuclear antisense RNA induces extensive adenosine modifications and nuclear retention of target transcripts
- PMID: 9108012
- PMCID: PMC20475
- DOI: 10.1073/pnas.94.8.3542
Nuclear antisense RNA induces extensive adenosine modifications and nuclear retention of target transcripts
Abstract
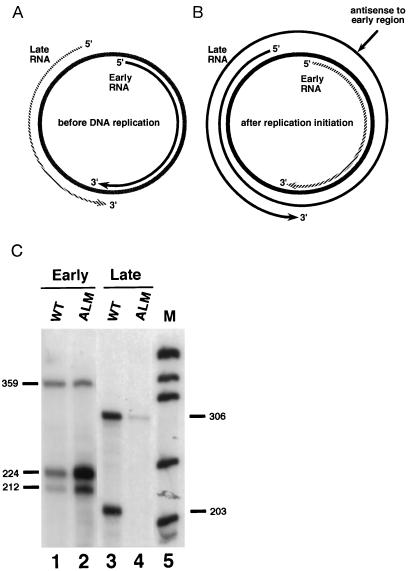
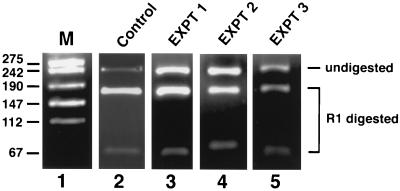
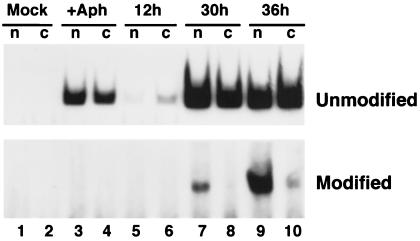
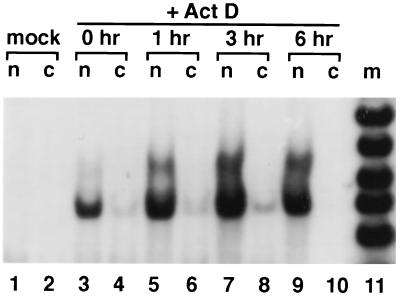
Antisense RNA may regulate the expression of a number of eukaryotic genes, but little is known about its prevalence or mechanism of action. We have used a model system in which antisense control can be studied both genetically and biochemically. Late in polyoma virus infection, early-strand mRNA levels are down-regulated by nuclear antisense RNA from the late strand. Analysis of early-strand transcripts isolated late in infection revealed extensive base modifications. In many transcripts almost half of the adenosines were altered to inosines or guanosines. These results suggest modification of RNA duplexes by double-stranded RNA adenosine deaminase or a related enzyme. Probes that detect only modified RNAs revealed that these molecules are not highly unstable, but accumulate within the nucleus and are thus inert for gene expression. Antisense-induced modifications can account for most or all of the observed regulation, with the lowered levels of early-strand RNAs commonly observed late in infection resulting from the fact that many transcripts are invisible to standard hybridization probes. This work suggests that similar antisense-mediated control mechanisms may also operate under physiological conditions in uninfected eukaryotic cells, and leads to the proposal that there is a novel pool of nuclear RNAs that cannot be seen with many molecular probes heretofore used.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
