

Independent, specialized invasions of ectomycorrhizal mutualism by two nonphotosynthetic orchids
- PMID: 9114020
- PMCID: PMC20753
- DOI: 10.1073/pnas.94.9.4510
Independent, specialized invasions of ectomycorrhizal mutualism by two nonphotosynthetic orchids
Abstract
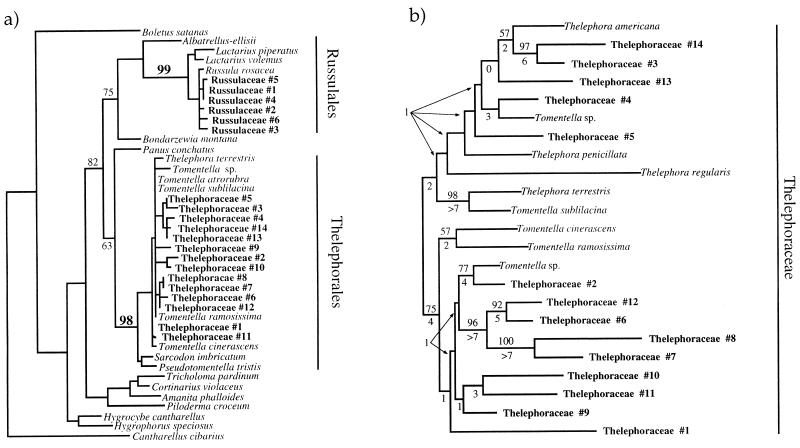
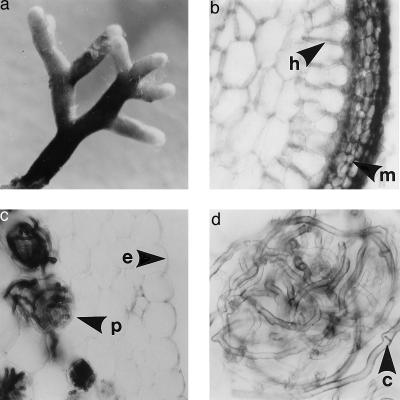
We have investigated the mycorrhizal associations of two nonphotosynthetic orchids from distant tribes within the Orchidaceae. The two orchids were found to associate exclusively with two distinct clades of ectomycorrhizal basidiomycetous fungi over wide geographic ranges. Yet both orchids retained the internal mycorrhizal structure typical of photosynthetic orchids that do not associate with ectomycorrhizal fungi. Restriction fragment length polymorphism and sequence analysis of two ribosomal regions along with fungal isolation provided congruent, independent evidence for the identities of the fungal symbionts. All 14 fungal entities that were associated with the orchid Cephalanthera austinae belonged to a clade within the Thelephoraceae, and all 18 fungal entities that were associated with the orchid Corallorhiza maculata fell within the Russulaceae. Restriction fragment length polymorphism and single-strand conformational polymorphism analysis of ectomycorrhizal tree roots collected adjacent to Cephalanthera showed that (i) the fungi associated internally with Cephalanthera also form typical external ectomycorrhizae and that (ii) ectomycorrhizae formed by other Basidiomycetes were abundant where the orchid grows but these fungi did not associate with the orchid. This is the first proof of ectomycorrhizal epiparasitism in nature by an orchid. We argue that these orchids are cheaters because they do not provide fixed carbon to associated fungi. This view suggests that mycorrhizae, like other ancient mutualisms, are susceptible to cheating. The extreme specificity in these orchids relative to other ectomycorrhizal plants agrees with trends seen in more conventional parasites.
Figures
References
-
- Mainero J S, Martinez del Rio C. In: The Biology of Mutualism: Ecology and Evolution. Boucher D H, editor. New York: Oxford Univ. Press; 1985. pp. 192–216.
-
- Boucher D H, James S, Keeler K H. Annu Rev Ecol Syst. 1982;13:315–347.
-
- Bronstein J L. Oikos. 1991;61:175–186.
-
- Pellmyr O, Leebens-Mack J, Huth C J. Nature (London) 1996;380:155–156. - PubMed
-
- Janzen D H. Science. 1975;188:936–937. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
