

Defective "pacemaker" current (Ih) in a zebrafish mutant with a slow heart rate
- PMID: 9114028
- PMCID: PMC20761
- DOI: 10.1073/pnas.94.9.4554
Defective "pacemaker" current (Ih) in a zebrafish mutant with a slow heart rate
Abstract
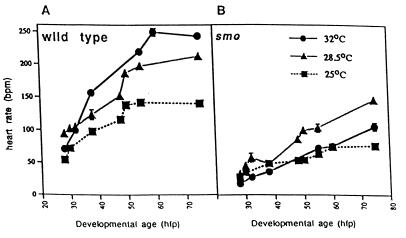
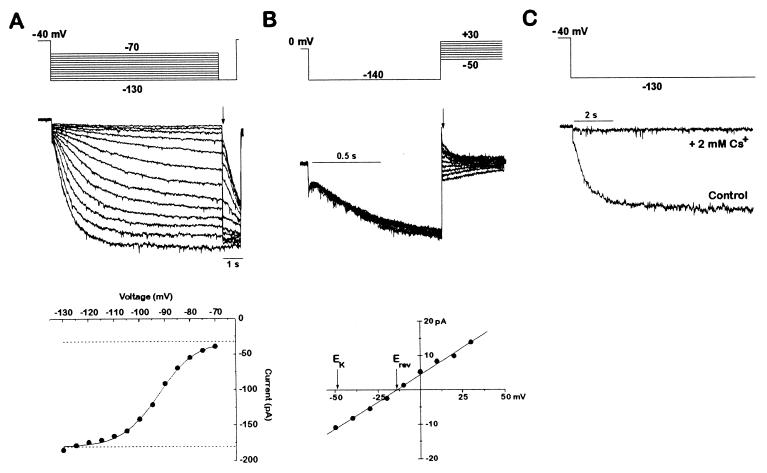
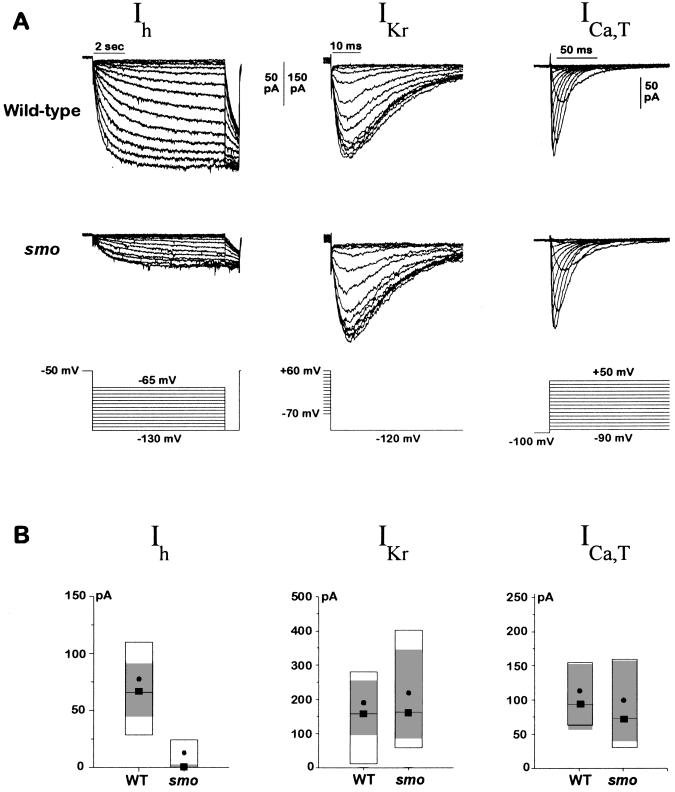
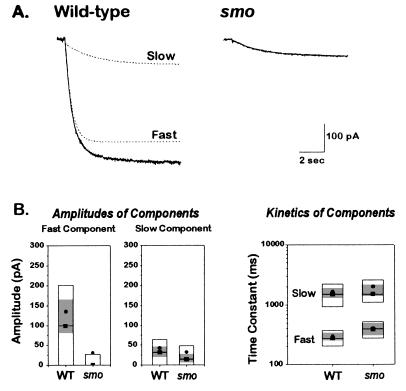
At a cellular level, cardiac pacemaking, which sets the rate and rhythm of the heartbeat, is produced by the slow membrane depolarization that occurs between action potentials. Several ionic currents could account for this pacemaker potential, but their relative prominence is controversial, and it is not known which ones actually play a pacemaking role in vivo. To correlate currents in individual heart cells with the rhythmic properties of the intact heart, we have examined slow mo (smo), a recessive mutation we discovered in the zebrafish Danio rerio. This mutation causes a reduced heart rate in the embryo, a property we can quantitate because the embryo is transparent. We developed methods for culture of cardiocytes from zebrafish embryos and found that, even in culture, cells from smo continue to beat relatively slowly. By patch-clamp analysis, we discovered that a large repertoire of cardiac currents noted in other species are present in these cultured cells, including sodium, T-type, and L-type calcium and several potassium currents, all of which appear normal in the mutant. The only abnormality appears to be in a hyperpolarization-activated inward current with the properties of Ih, a current described previously in the nervous system, pacemaker, and other cardiac tissue. smo cardiomyocytes have a reduction in Ih that appears to result from severe diminution of one kinetic component of the Ih current. This provides strong evidence that Ih is an important contributor to the pacemaking behavior of the intact heart.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
