

Hedgehog signaling regulates transcription through cubitus interruptus, a sequence-specific DNA binding protein
- PMID: 9122207
- PMCID: PMC20100
- DOI: 10.1073/pnas.94.6.2404
Hedgehog signaling regulates transcription through cubitus interruptus, a sequence-specific DNA binding protein
Abstract
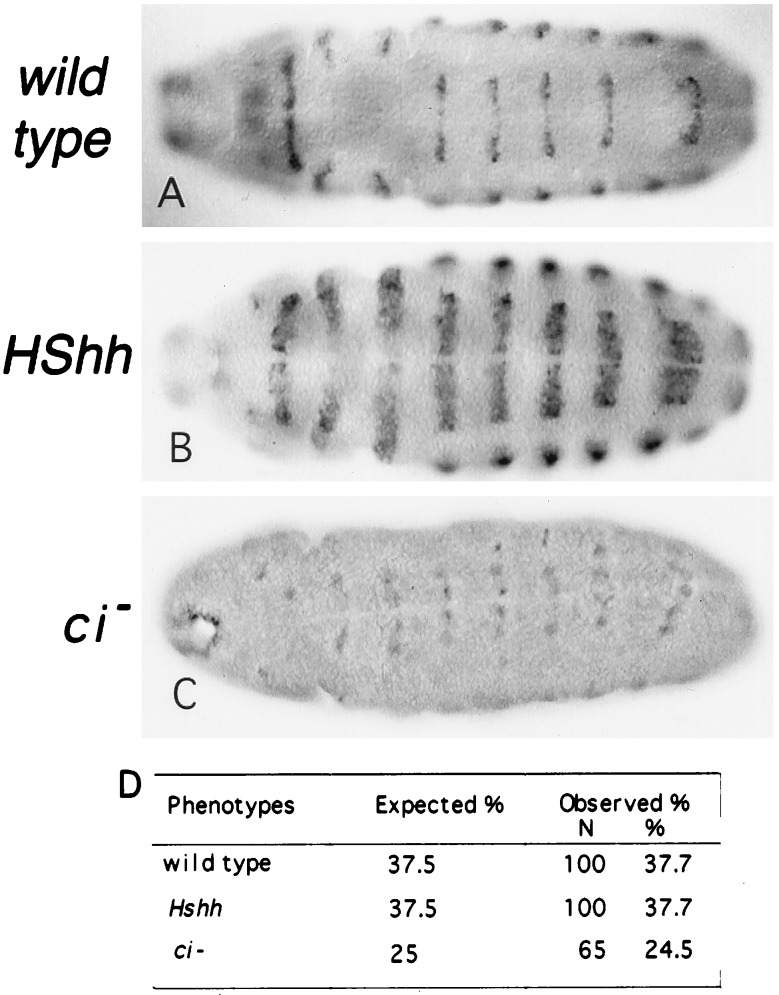
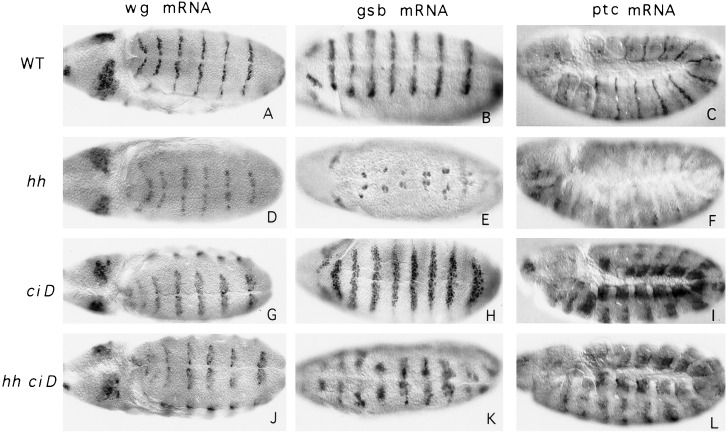
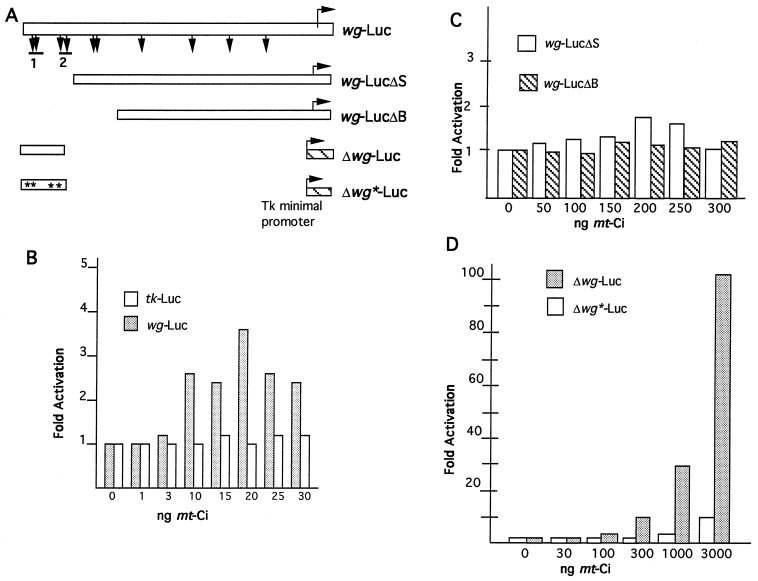
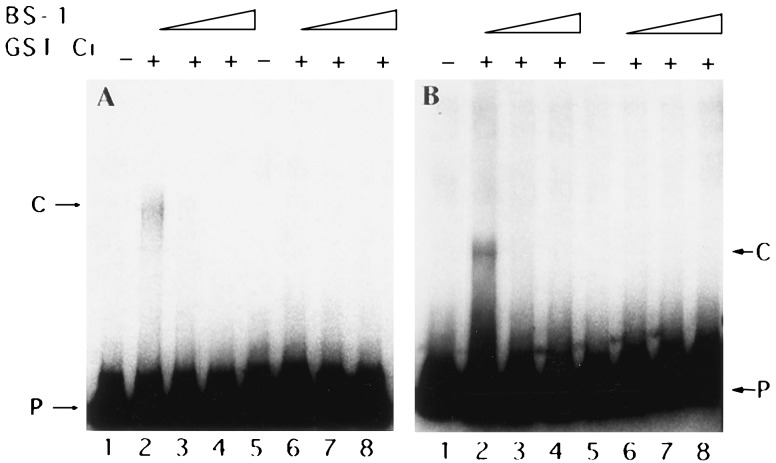
Hedgehog (Hh) is a member of a family of secreted proteins that direct patterning at multiple stages in both Drosophila and vertebrate development. During Drosophila embryogenesis, Hh protein is secreted by the cells of the posterior compartment of each segment. hh activates transcription of wingless (wg), gooseberry (gsb), and patched (ptc) in the cells immediately adjacent to Hh-secreting cells. Hh signaling is thought to involve the segment polarity gene cubitus interruptus (ci). ci encodes a zinc finger protein of the Gli family of sequence-specific DNA binding proteins. ci mRNA is expressed in all non-Hh expressing cells. Here we demonstrate ci activity is both necessary and sufficient to drive expression of Hh-responsive genes in the Drosophila embryos. We show that Ci is a sequence-specific DNA binding protein that drives transcription from the wg promoter in transiently transfected cells. We demonstrate that Ci binding sites in the wg promoter are necessary for this transcriptional activation. These data taken together provide strong evidence that Ci is a transcriptional effector of Hh signaling.
Figures

References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
