

doi: 10.1073/pnas.94.6.2545.
DNA methylation and genetic instability in colorectal cancer cells
Affiliations
- PMID: 9122232
- PMCID: PMC20125
- DOI: 10.1073/pnas.94.6.2545
Item in Clipboard
DNA methylation and genetic instability in colorectal cancer cells
Proc Natl Acad Sci U S A.
.
Abstract
Apparent alterations in DNA methylation have been observed in many cancers, but whether such alterations represent a persistent alteration in the normal methylation process is not known. In this study, we report a striking difference in the expression of exogenously introduced retroviral genes in various colorectal cancer cell lines. Extinguished expression was associated with DNA methylation and could be reversed by treatment with the demethylating agent 5-azacytidine. A striking correlation between genetic instability and methylation capacity suggested that methylation abnormalities may play a role in chromosome segregation processes in cancer cells.
Figures
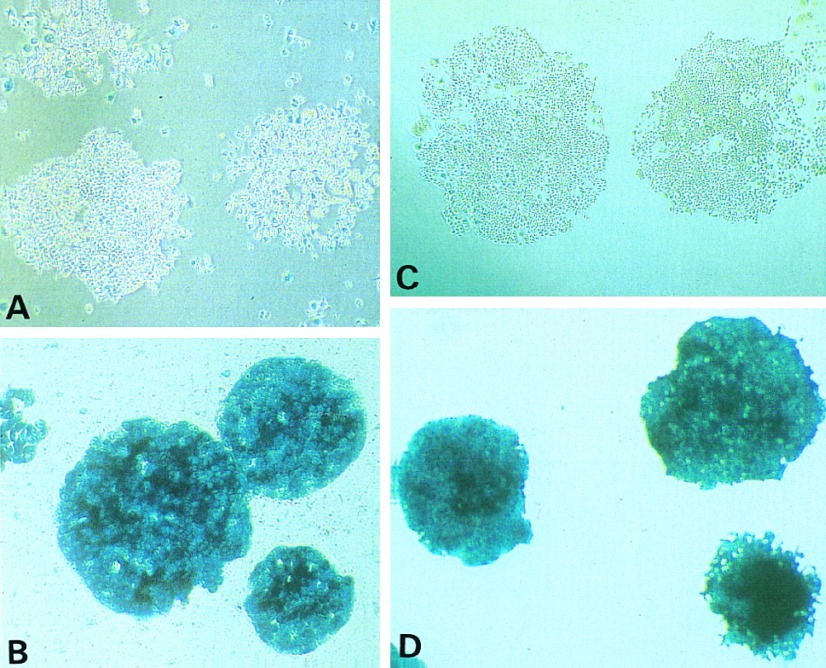
Histochemical detection of β-gal expression. (A and B) After infection with the retrovirus Gal/Neo and selection with G418, cells were stained in situ with X-Gal to confirm the expression of β-gal. Whereas the majority of cells from individual clones of cell line HCT116 was not stained (A), virtually all cells of HT29 stained intensely blue (B). (C and D) Cells were stained in situ with X-Gal after retroviral infection with Gal/Hyg and selection with hygromycin. Whereas virtually all HT29 cells stained blue (D), the majority of cells from individual clones of HCT116+ch3 were not stained (C). (All ×10.)
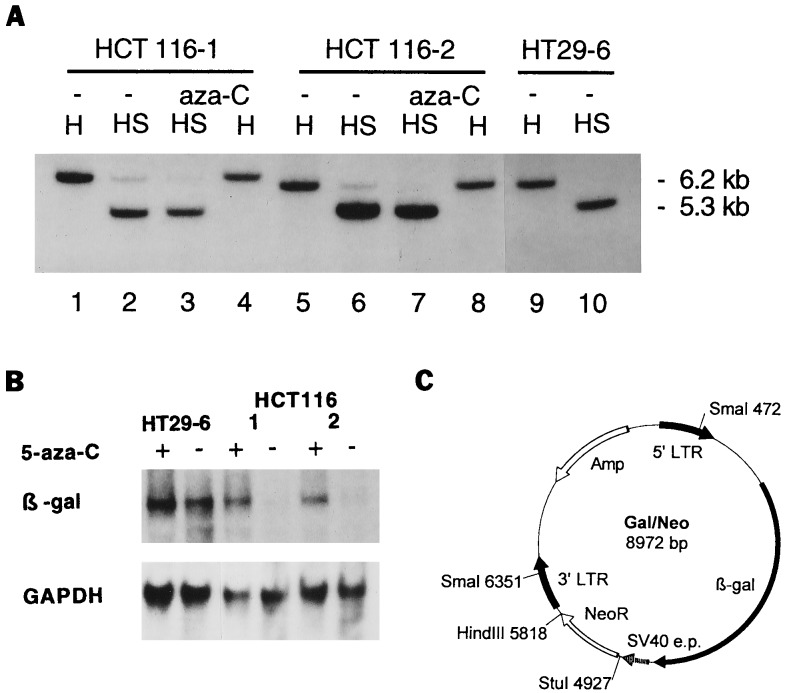
Methylation of β-gal gene after retroviral infection. (A) Analysis of 5′LTR promoter by methylation-sensitive restriction enzyme. DNA from untreated (−) or 5-aza-C-treated cells was digested with HindIII alone (H) or with HindIII plus SmaI (HS). The blotted gel was probed with a [32P]dCTP-labeled fragment of the β-gal gene. The HindIII fragment containing β-gal sequences was 6.2 kb in clone HT29–6 (lane 9), and SmaI reduced this fragment to 5.35 kb (lane 10), reflecting the invariant size separating the HindIII and SmaI sites in the provirus Gal/Neo. The SmaI digest of the HCT116 clones (lane 2 and 6) revealed two bands, indicating methylation of the SmaI site in these cells. Treatment of the HCT116 clones with 5-aza-C resulted in the removal of methyl groups, so that only a 5.35-kb fragment was detected after SmaI plus HindIII digestion (lanes 3 and 7). (B) Transcription of β-gal gene. Total RNA of the clones HCT116–1, HCT116–2, and HT29–6 was probed with a [32P]dCTP-labeled fragment of the β-gal gene. The level of expression of β-gal mRNA in clones HCT116–1 and 2 was low (lanes 4 and 6) but increased substantially after 5-aza-C treatment (lanes 3 and 5). The 5-aza-C treatment had no effect on β-gal mRNA of clone HT29–6 (lanes 1 and 2). Rehybridization of the filter with a glyceraldehyde-3-phosphate dehydrogenase (GAPDH) probe confirmed that equal amounts of RNA were loaded. (C) Scheme of retroviral vector Gal/Neo. The Gal/Neo vector contains the β-gal reporter driven by a retroviral LTR and the G418-resistance gene (NeoR) under the control of the simian virus 40 early promoter (SV40 e.p.). Positions of the two SmaI and the single HindIII restriction sites are indicated. The restriction sites HindIII and StuI were used for the construction of the provirus Gal/Hyg.
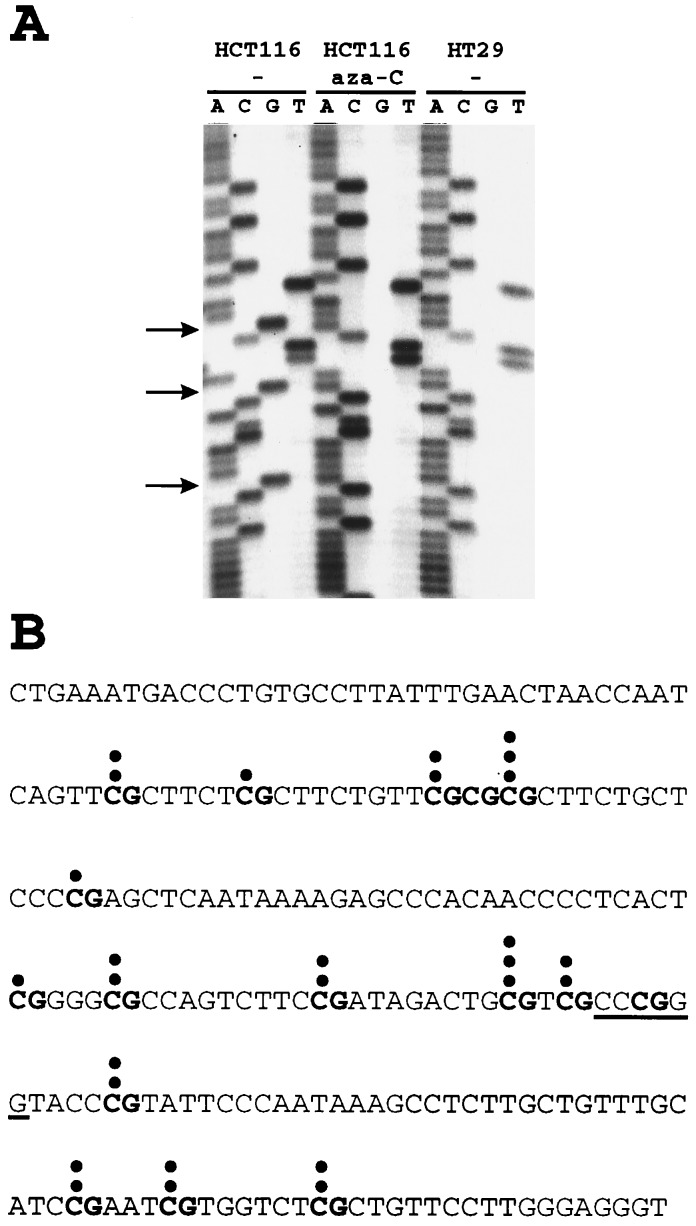
Bisulfite genomic sequencing of 5′LTR. Genomic DNAs of infected cell lines were treated with bisulfite, converting deoxycytosine but not 5-methylcytosine residues into uracil through deamination. After treatment, the viral 5′LTR region was PCR-amplified, the PCR products were cloned, and a 214-bp region surrounding the SmaI site (see Fig. 2C) was sequenced. (A) Examples of clones HCT116–1 (before and after 5-aza-C treatment) and HT29–6 are shown. Arrows mark the locations of methylated CpG dinucleotides in HCT116–1. (B) Distribution of methylated CpG dinucleotides within the 5′LTR promotor of HCT116–1. The DNA sequence of the 214-bp region surrounding the SmaI site (underlined) is shown. Its 16 CpG dinucleotides are typed in boldface. The shaded area represents the region that is shown on the sequencing gel in A. Each dot represents a 5-methylcytosine residue that was not converted into uracil in one of the 12 cloned PCR products.
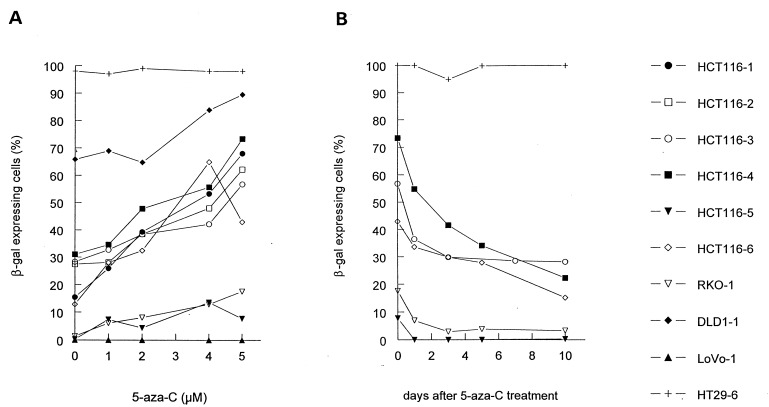
Reversal of β-gal inactivation by 5-aza-C. (A) Nine MMR-deficient clones (HCT116–1, HCT116–2, HCT116–3, HCT116–4, HCT116–5, HCT116–6, RKO-1, DLD1–1, and LoVo-1) and the MMR-proficient clone HT29–6 were treated with different concentrations (0, 1, 2, 4, or 5 μM) of 5-aza-C for 72 hr and stained with X-Gal. In 8 of 9 MMR-deficient clones, 5-aza-C significantly increased the fraction of cells expressing β-gal in a dose-dependent manner. The single MMR-deficient clone in which 5-aza-C had no effect was LoVo-1. LoVo-1 had a truncating mutation in the β-gal gene (see text). (B) To determine whether the reversal of inactivation of β-gal expression by 5-aza-C was permanent, the drug (5 μM) was removed from the medium, and β-gal expression was measured at subsequent times. The clones reverted to the same basal level of β-gal expression found before 5-aza-C treatment.
Comment in
-
Altered DNA methylation and genome instability: a new pathway to cancer?Proc Natl Acad Sci U S A. 1997 Mar 18;94(6):2103-5. doi: 10.1073/pnas.94.6.2103. Proc Natl Acad Sci U S A. 1997. PMID: 9122155 Free PMC article. Review. No abstract available.
References
-
- Bird A. Cell. 1992;70:5–8. - PubMed
-
- Noyer-Weldner M, Trautner T A. In: DNA Methylation: Molecular Biology and Biological Significance. Jost J P, Saluz H P, editors. Basel: Birkhauser; 1993. pp. 39–108.
-
- Yeivin A, Razin A. In: DNA Methylation: Molecular Biology and Biological Significance. Jost J P, Saluz H P, editors. Basel: Birkhauser; 1993. pp. 523–568.
-
- Bird A. Nature (London) 1986;321:209–213. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
