

The Saccharomyces cerevisiae kinesin-related motor Kar3p acts at preanaphase spindle poles to limit the number and length of cytoplasmic microtubules
- PMID: 9128252
- PMCID: PMC2139775
- DOI: 10.1083/jcb.137.2.417
The Saccharomyces cerevisiae kinesin-related motor Kar3p acts at preanaphase spindle poles to limit the number and length of cytoplasmic microtubules
Abstract
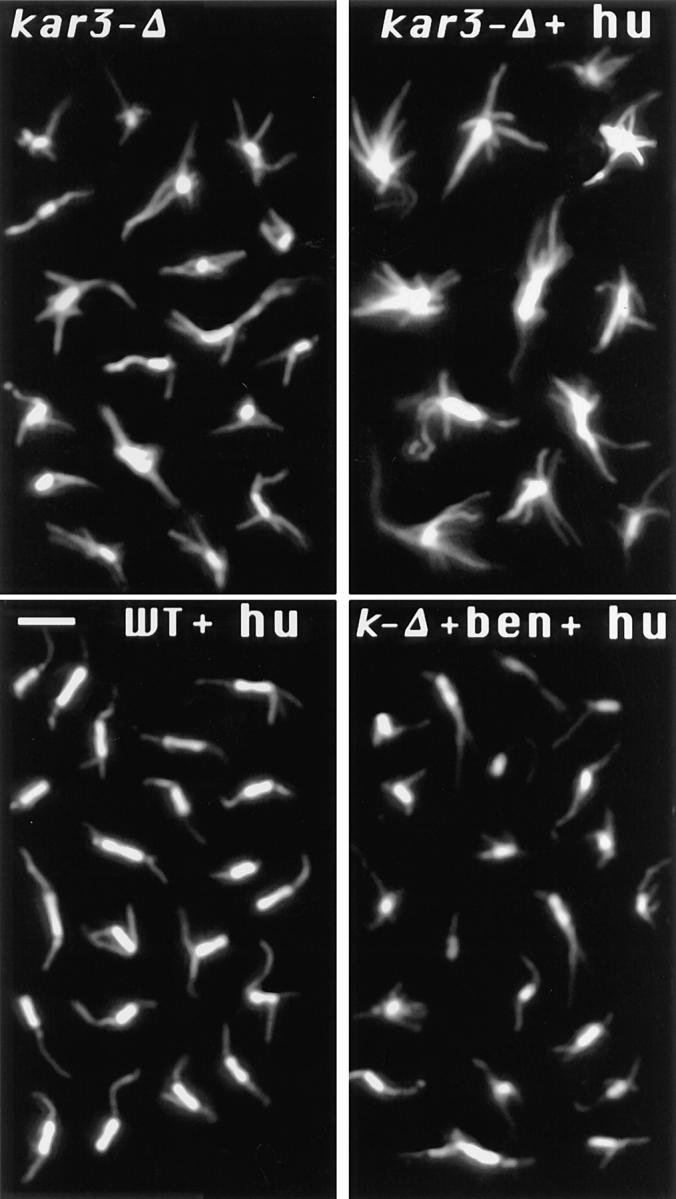
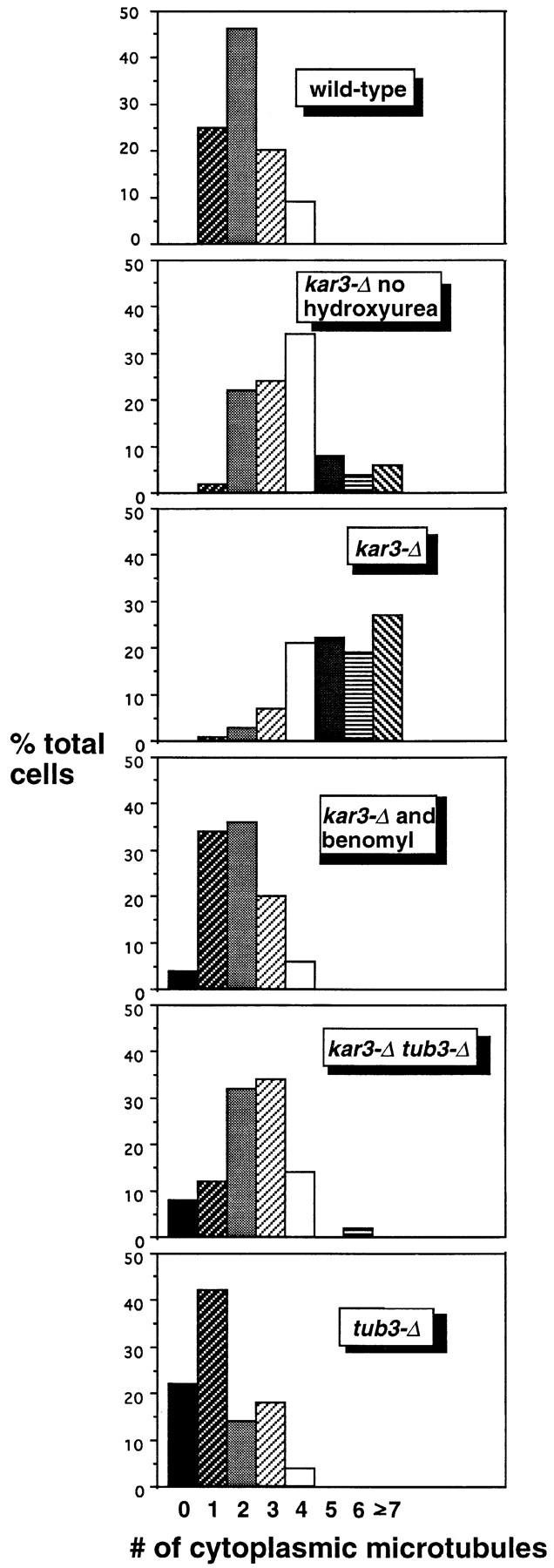
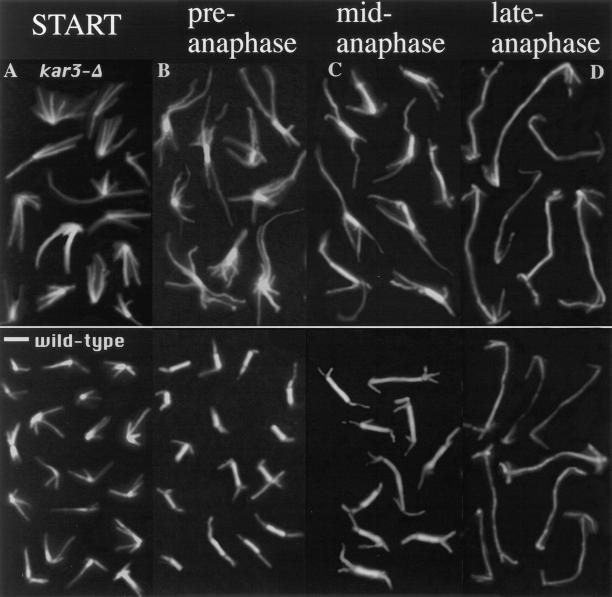
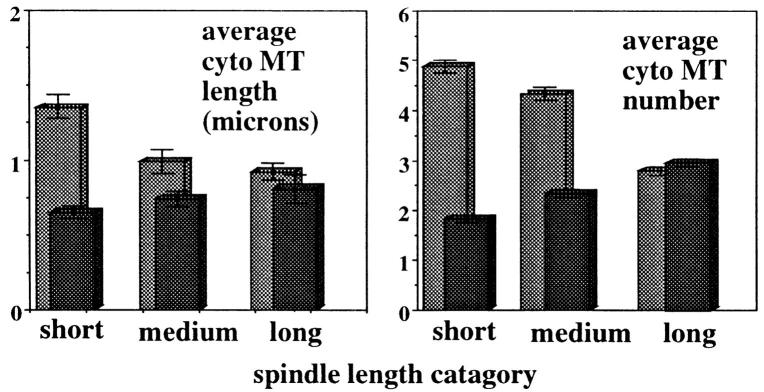
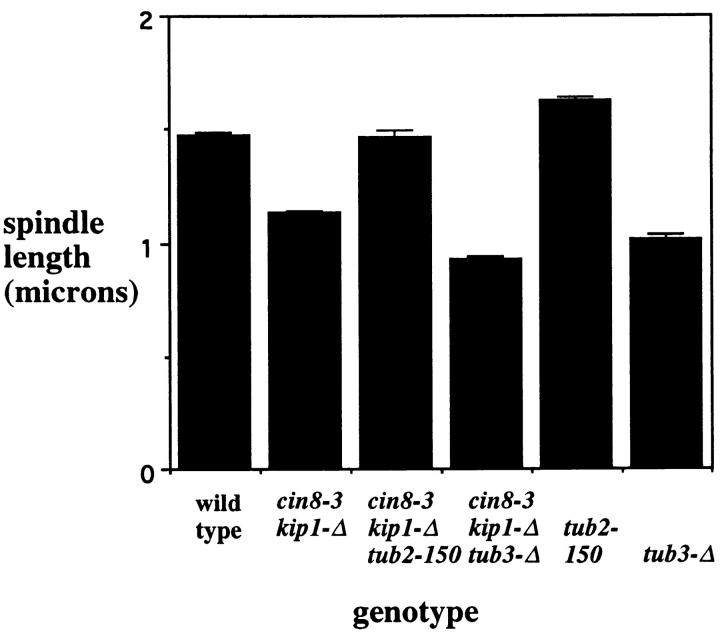
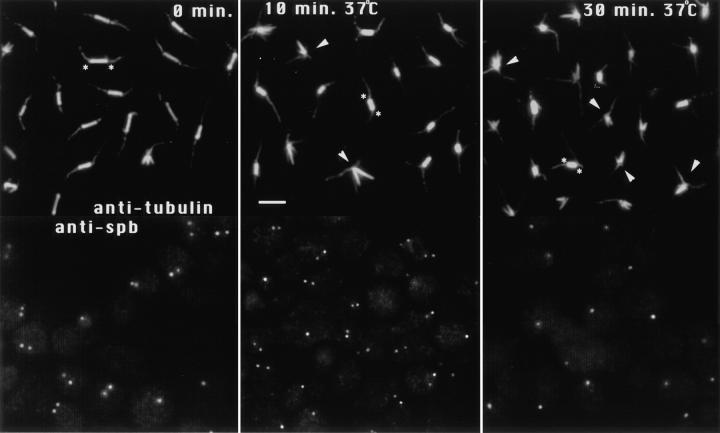
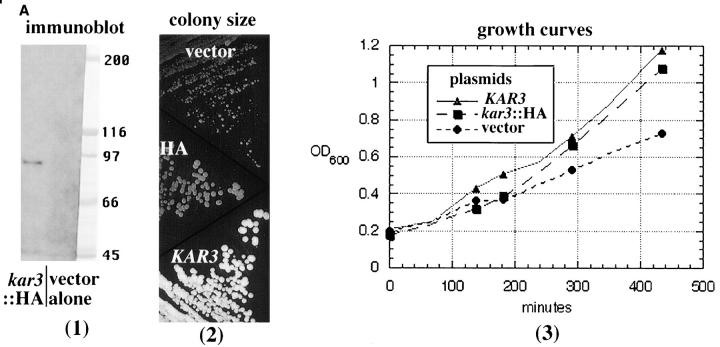
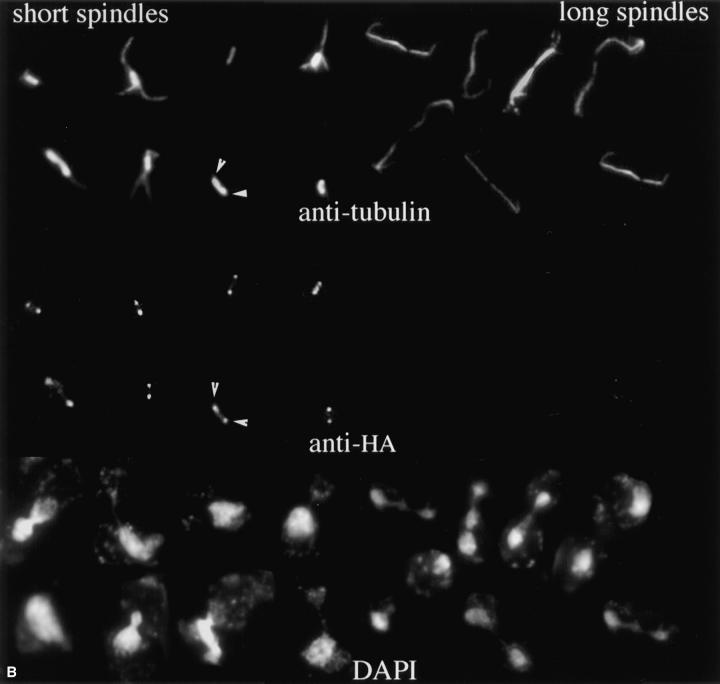
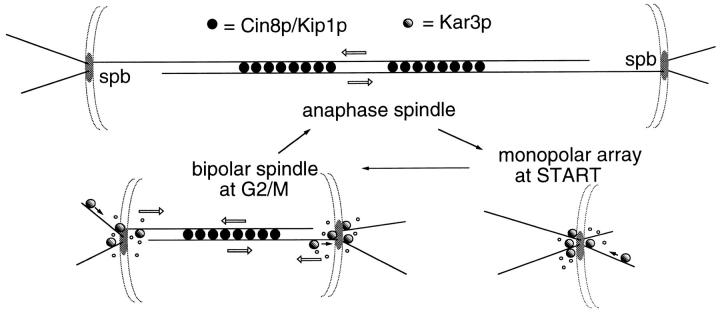
The Saccharomyces cerevisiae kinesin-related motor Kar3p, though known to be required for karyogamy, plays a poorly defined, nonessential role during vegetative growth. We have found evidence suggesting that Kar3p functions to limit the number and length of cytoplasmic microtubules in a cell cycle-specific manner. Deletion of KAR3 leads to a dramatic increase in cytoplasmic microtubules, a phenotype which is most pronounced from START through the onset of anaphase but less so during late anaphase in synchronized cultures. We have immunolocalized HA-tagged Kar3p to the spindle pole body region, and fittingly, Kar3p was not detected by late anaphase. A microtubule depolymerizing activity may be the major vegetative role for Kar3p. Addition of the microtubule polymerization inhibitors nocodazol or benomyl to the medium or deletion of the nonessential alpha-tubulin TUB3 gene can mostly correct the abnormal microtubule arrays and other growth defects of kar3 mutants, suggesting that these phenotypes result from excessive microtubule polymerization. Microtubule depolymerization may also be the mechanism by which Kar3p acts in opposition to the anaphase B motors Cin8p and Kip1p. A preanaphase spindle collapse phenotype of cin8 kip1 mutants, previously shown to involve Kar3p, is markedly delayed when microtubule depolymerization is inhibited by the tub2-150 mutation. These results suggest that the Kar3p motor may act to regulate the length and number of microtubules in the preanaphase spindle.
Figures

References
-
- Belmont LD, Hyman AA, Sawin KE, Mitchison TJ. Real-time visualization of cell cycle dependent changes in microtubule dynamics in cytoplasmic extracts. Cell. 1990;62:579–589. - PubMed
-
- Davidse LC. Benzimadazole fungicides: mechanism of action and biological impact. Annu Rev Phytopathol. 1986;24:43–65.
-
- De Brabander M. Microtubule dynamics during the cell cycle: the effects of taxol and nocodazole on the microtubule system of PtK2 cells at different stages of the mitotic cycle. Int Rev Cytol. 1986;101:215–274. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
