

Arachidonic acid inhibits transient potassium currents and broadens action potentials during electrographic seizures in hippocampal pyramidal and inhibitory interneurons
- PMID: 9133373
- PMCID: PMC6573692
- DOI: 10.1523/JNEUROSCI.17-10-03476.1997
Arachidonic acid inhibits transient potassium currents and broadens action potentials during electrographic seizures in hippocampal pyramidal and inhibitory interneurons
Abstract
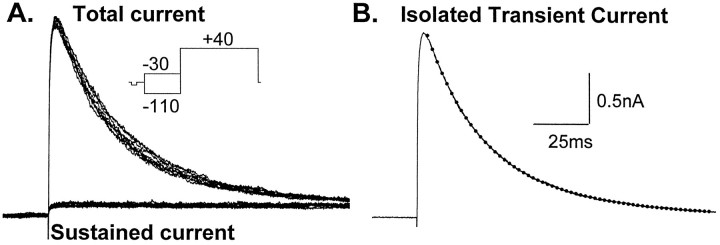
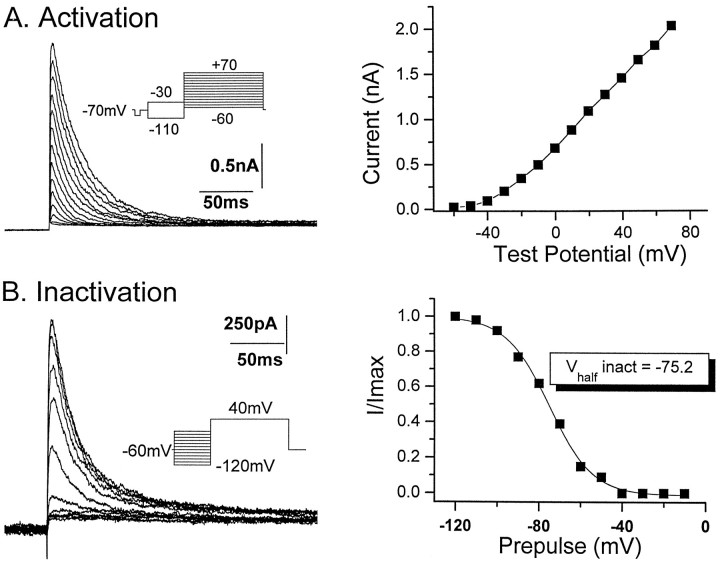
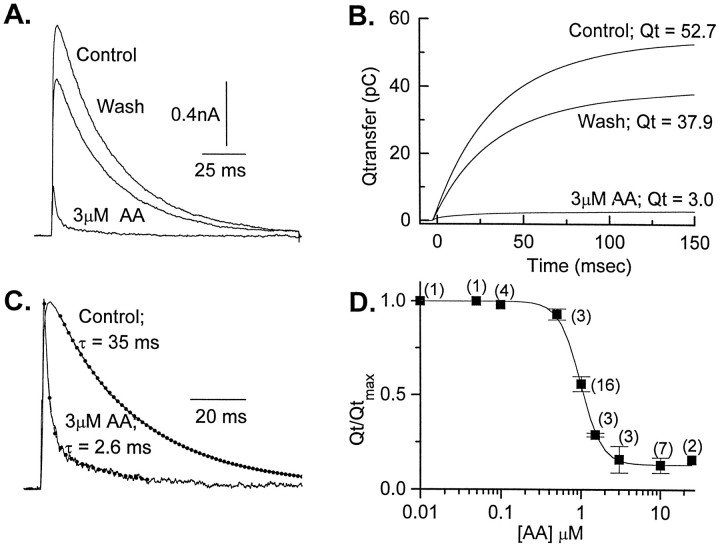
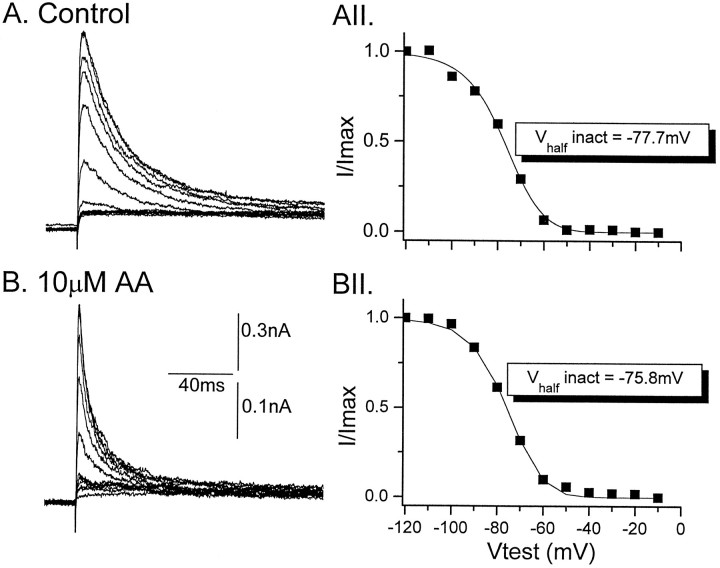
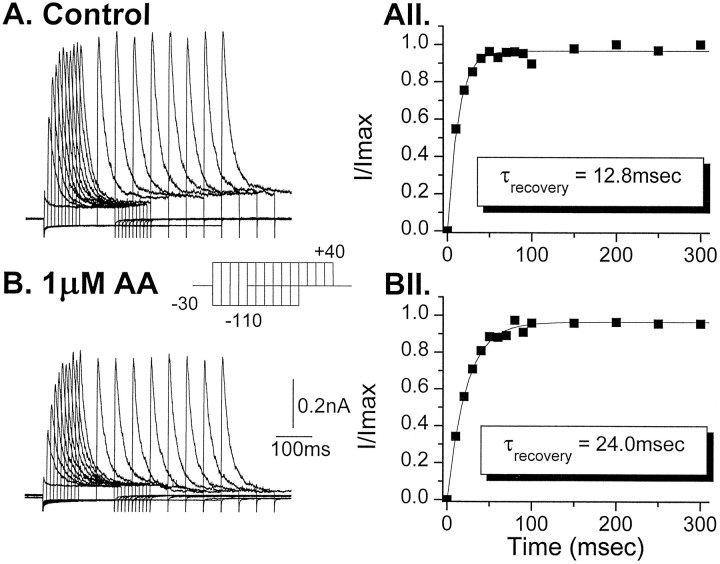
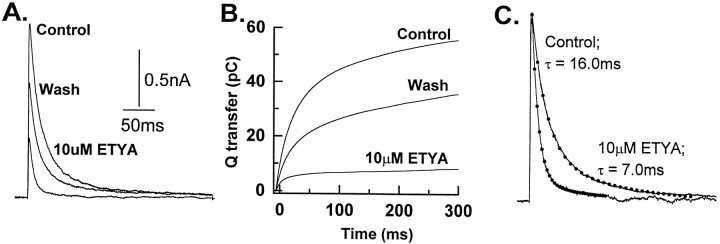
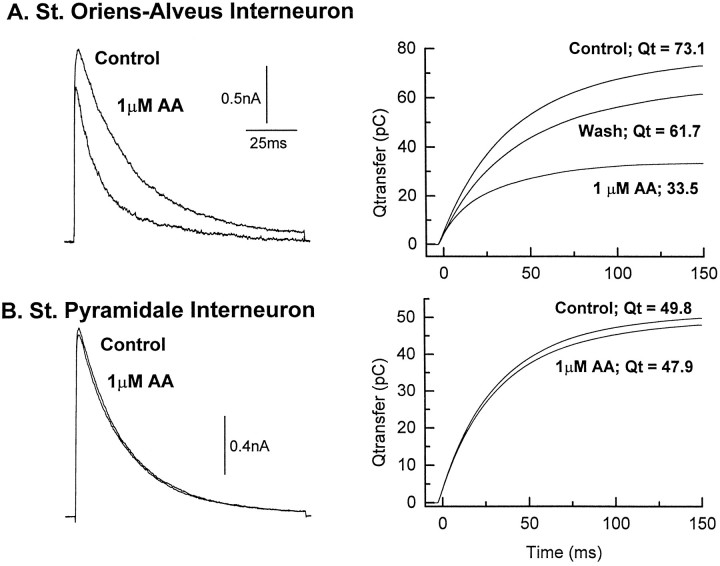
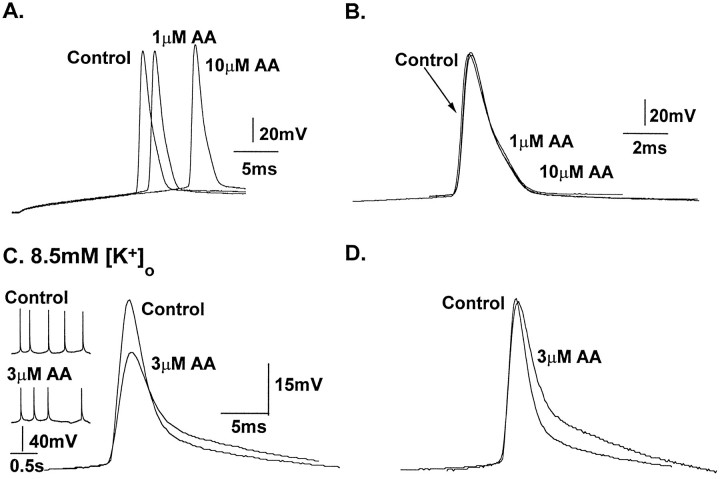
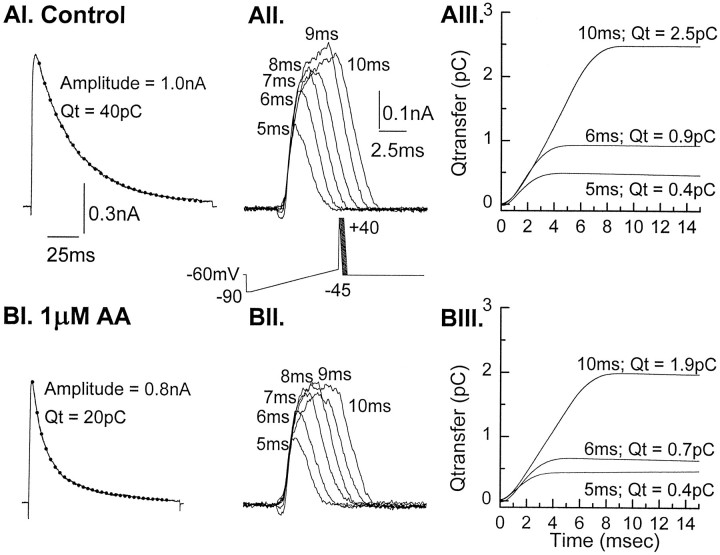
The transient outward potassium current was studied in outside-out macropatches excised from the soma of CA1 pyramidal neurons and stratum (st.) oriens-alveus inhibitory interneurons in rat hippocampal slices. Arachidonic acid dose dependently decreased the charge transfer associated with the transient current, concomitant with an increase in the rate of current inactivation. Arachidonic acid (AA) did not affect the voltage dependence of steady state inactivation but did prolong the period required for complete recovery from inactivation. The effects of AA were mimicked by the nonmetabolizable analog of AA, 5,8,11,14-eicosatetraynoic acid, suggesting that metabolic products of AA were not responsible for the observed blocking action. In addition, AA blocked st. oriens-alveus-lacunosum-moleculare interneuron transient currents but not currents recorded from basket cell interneurons. In current clamp experiments, AA was without effect on the action potential waveform of CA1 pyramidal neurons under control recording conditions. In voltage-clamp experiments, the use of a test pulse paradigm, designed to mimic the action potential voltage trajectory, revealed that the transient current normally associated with a single spike deactivates too rapidly for AA to have an effect. Transient currents activated by longer duration "action potential" waveforms, however, were attenuated by AA. Consistent with this finding was the observation that AA broadened interictal spikes recorded in the elevated [K+]o model of epilepsy. These data suggest that AA liberated from hippocampal neurons may act to block the transient current selectively in both CA1 pyramidal neurons and inhibitory interneurons and to broaden action potentials selectively under pathological conditions.
Figures
References
-
- Axelrod J. Receptor-mediated activation of phospholipase A2 and arachidonic acid release in signal transduction. Biochem Soc Trans. 1990;18:503–507. - PubMed
-
- Bazan NG, Birkle DL, Tang W, Reddy TS. The accumulation of free arachidonic acid, diacylglycerols, prostaglandins, and lipoxygenase reaction products in the brain during experimental epilepsy. In: Degado-Escueta AV, Ward AA Jr, Woodbury DM, Porter RJ, editors. Advances in neurology, Ed 44. Raven; New York: 1986. pp. 879–902. - PubMed
-
- Bevan S, Wood JN. Arachidonic-acid metabolites as second messengers. Nature. 1987;328:20–22. - PubMed
-
- Bito H, Mori M, Sakanaka C, Takano T, Zen-ichiro H, Gotoh Y, Nishida E, Shimizu T. Functional coupling of the SSTR4, a major hippocampal somatostatin receptor, to adenylate cyclase inhibition, arachidonate release, and activation of the mitogen-activated protein kinase cascade. J Biol Chem. 1993;269:12722–12730. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous