

Localized and transient elevations of intracellular Ca2+ induce the dedifferentiation of axonal segments into growth cones
- PMID: 9133380
- PMCID: PMC6573697
- DOI: 10.1523/JNEUROSCI.17-10-03568.1997
Localized and transient elevations of intracellular Ca2+ induce the dedifferentiation of axonal segments into growth cones
Abstract
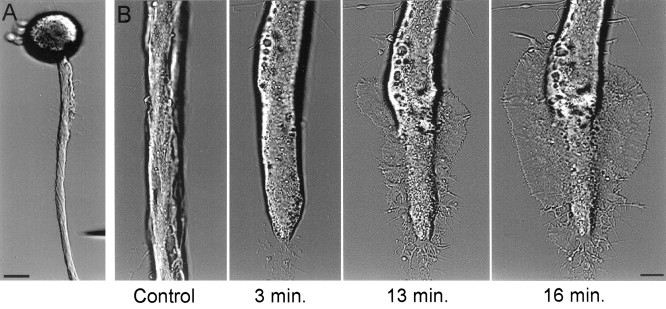
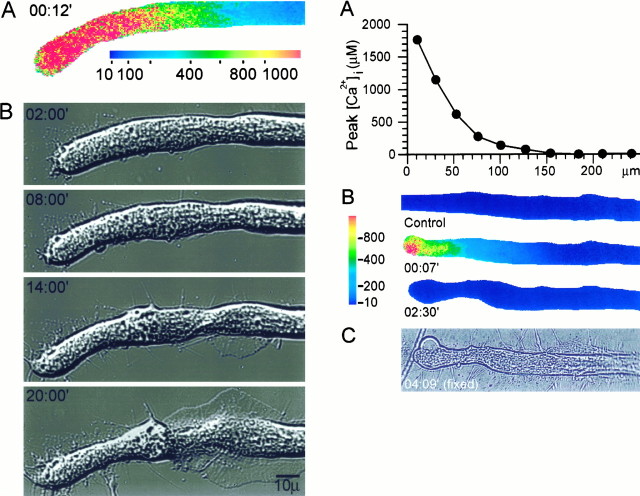
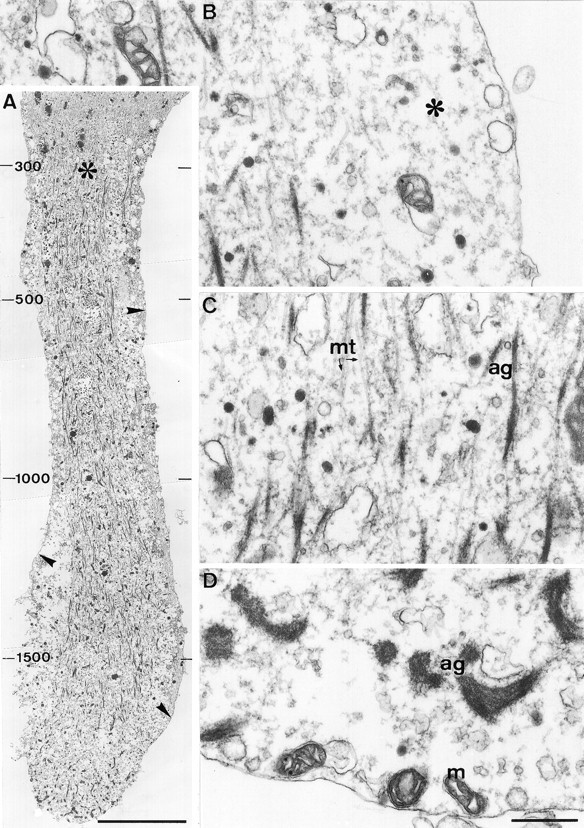
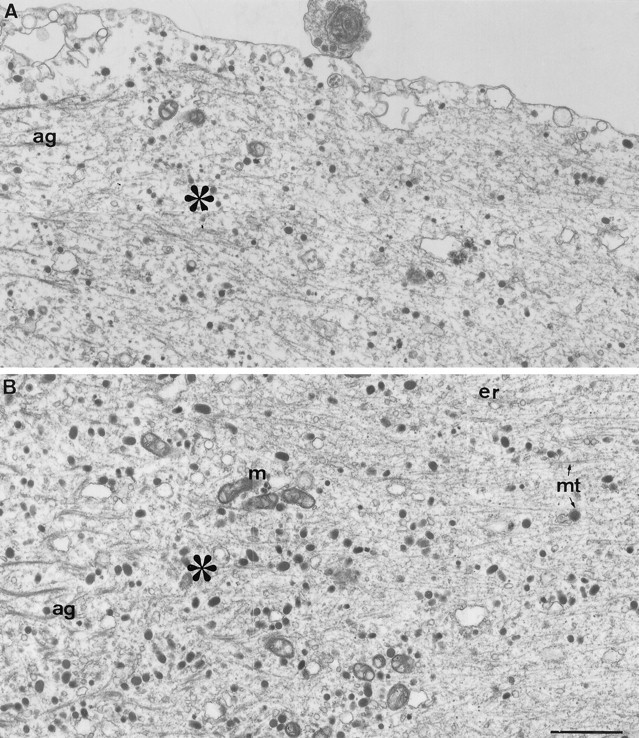
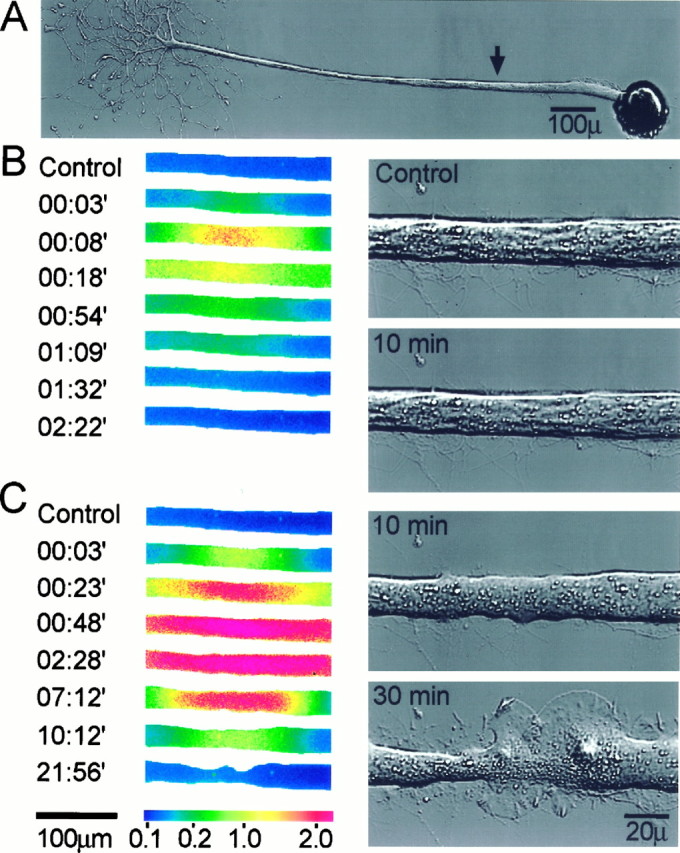
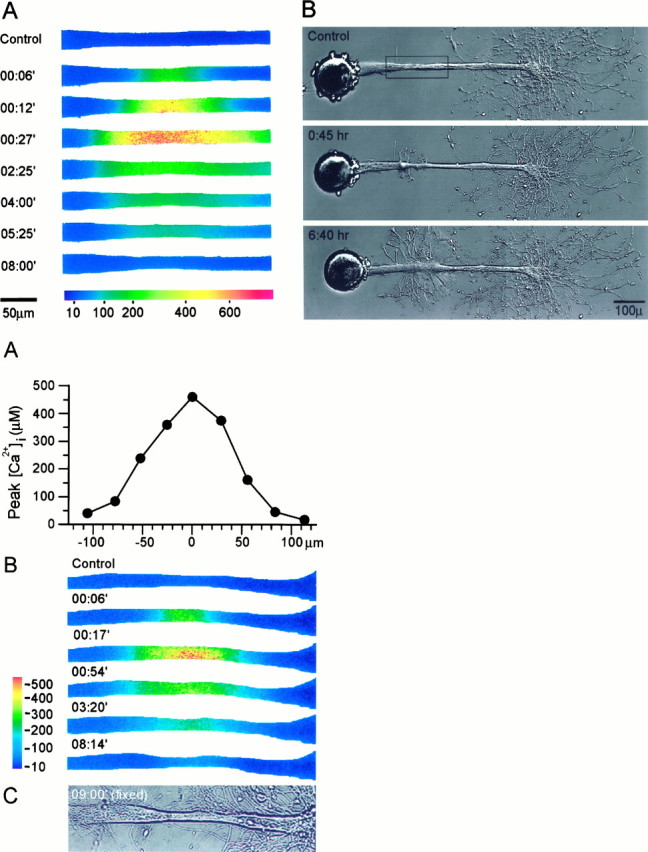
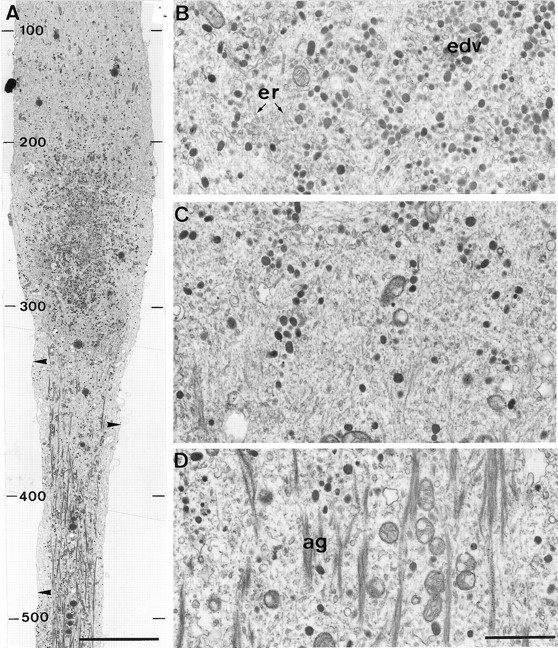
The formation of a growth cone at the tip of a severed axon is a key step in its successful regeneration. This process involves major structural and functional alterations in the formerly differentiated axonal segment. Here we examined the hypothesis that the large, localized, and transient elevation in the free intracellular calcium concentration ([Ca2+]i) that follows axotomy provides a signal sufficient to trigger the dedifferentiation of the axonal segment into a growth cone. Ratiometric fluorescence microscopy and electron microscopy were used to study the relations among spatiotemporal changes in [Ca2+]i, growth cone formation, and ultrastructural alterations in axotomized and intact Aplysia californica neurons in culture. We report that, in neurons primed to grow, a growth cone forms within 10 min of axotomy near the tip of the transected axon. The nascent growth cone extends initially from a region in which peak intracellular Ca2+ concentrations of 300-500 microM are recorded after axotomy. Similar [Ca2+]i transients, produced in intact axons by focal applications of ionomycin, induce the formation of ectopic growth cones and subsequent neuritogenesis. Electron microscopy analysis reveals that the ultrastructural alterations associated with axotomy and ionomycin-induced growth cone formation are practically identical. In both cases, growth cones extend from regions in which sharp transitions are observed between axoplasm with major ultrastructural alterations and axoplasm in which the ultrastructure is unaltered. These findings suggest that transient elevations of [Ca2+]i to 300-500 microM, such as those caused by mechanical injury, may be sufficient to induce the transformation of differentiated axonal segments into growth cones.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous