

Failed cell migration and death of purkinje cells and deep nuclear neurons in the weaver cerebellum
- PMID: 9133390
- PMCID: PMC6573684
- DOI: 10.1523/JNEUROSCI.17-10-03675.1997
Failed cell migration and death of purkinje cells and deep nuclear neurons in the weaver cerebellum
Abstract
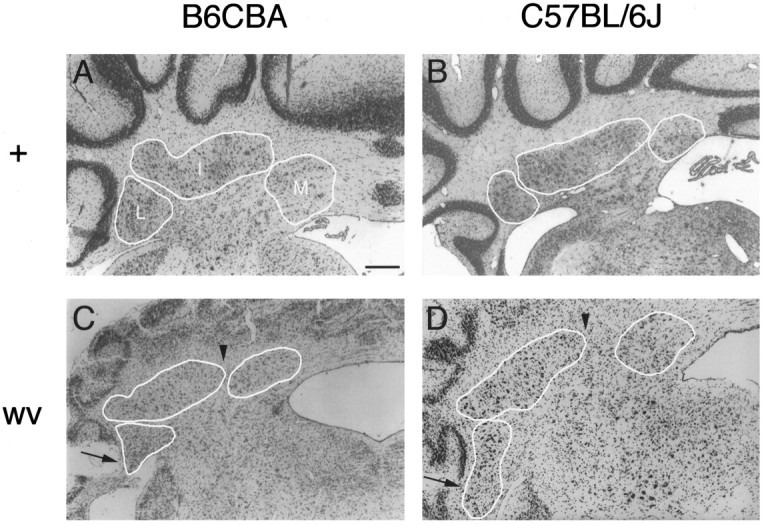
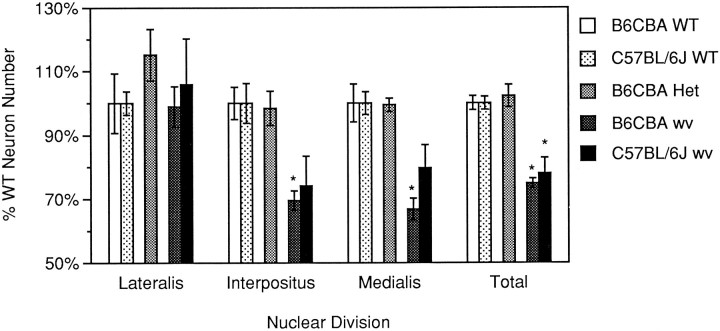
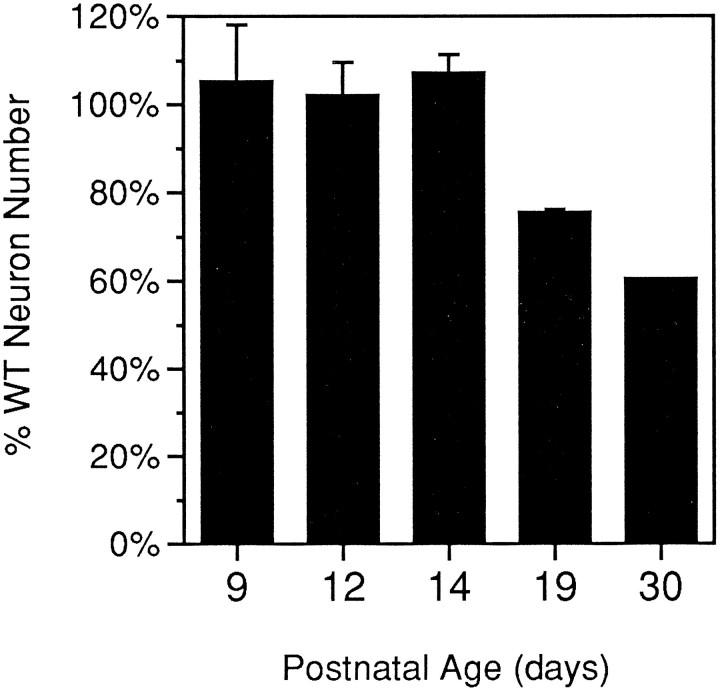
The mouse neurological mutant weaver has an atrophic cerebellar cortex with deficits in both Purkinje and granule cell number. Although granule cells are known to die postnatally shortly after their final cell division, the cause of the Purkinje cell deficit (cell death vs lack of production) is unknown. We report here a quantitative analysis of large cerebellar neurons of the weaver mutant during postnatal development. We explored the hypothesis that the cells of the entire cerebellar anlage were affected by the mutation by including in our study the neurons of the deep cerebellar nuclei (DCN). Our analysis reveals that in homozygous weaver mutants (1) the DCN are displaced laterally, display an abnormal anatomy, and suffer a 20-25% decrease in neuron number; (2) this numerical deficit is located in medial regions, similar to the localization of cortical deficits in both Purkinje and granule cells; (3) pyknotic figures are present in the juvenile DCN and in the Purkinje cell layer; and (4) the majority of cell death in these populations occurs not in medial regions where the numerical deficits are observed, but rather laterally where adult cell number is nearly normal. These results lead us to propose that the complete weaver phenotype includes a failure of the cell movements that lead to the fusion of the bilateral cerebellar anlage, and that this failure to migrate properly leaves some of the Purkinje cells and DCN neurons in a position where they are unable to make appropriate connections, leading to their death. In addition to implications for normal development, these observations suggest that weaver effects on the cerebellum can be unified into one consolidated model in which failure of cell movement affects all major cerebellar neurons.
Figures




References
-
- Altman J, Anderson WJ. Experimental reorganization of the cerebellar cortex. I. Morphological effects of elimination of all microneurons with prolonged x-irradiation started at birth. J Comp Neurol. 1972;146:355–406. - PubMed
-
- Altman J, Bayer S. Embryonic development of the rat cerebellum. I. Delineation of the cerebellar primordium and early cell movements. J Comp Neurol. 1985a;231:1–26. - PubMed
-
- Altman J, Bayer S. Embryonic development of the rat cerebellum. II. Translocation and regional distribution of the deep neurons. J Comp Neurol. 1985b;231:27–41. - PubMed
-
- Altman J, Bayer S. Embryonic development of the rat cerebellum. III. Regional differences in the time of origin, migration and settling of Purkinje cells. J Comp Neurol. 1985c;231:42–65. - PubMed
-
- Armstrong DM, Schild RF. An investigation of the cerebellar cortico-nuclear projections in the rat using an autoradiographic tracing method. I. Projection from the vermis. Brain Res. 1978a;141:1–19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous