

Dentate granule cell neurogenesis is increased by seizures and contributes to aberrant network reorganization in the adult rat hippocampus
- PMID: 9133393
- PMCID: PMC6573703
- DOI: 10.1523/JNEUROSCI.17-10-03727.1997
Dentate granule cell neurogenesis is increased by seizures and contributes to aberrant network reorganization in the adult rat hippocampus
Abstract
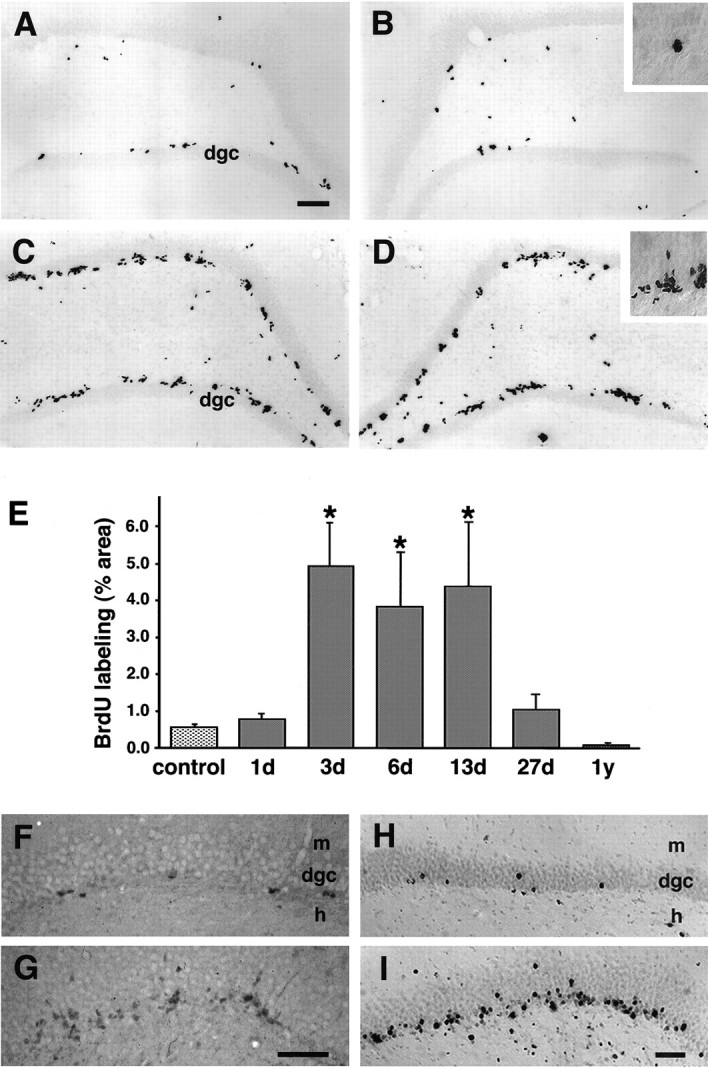
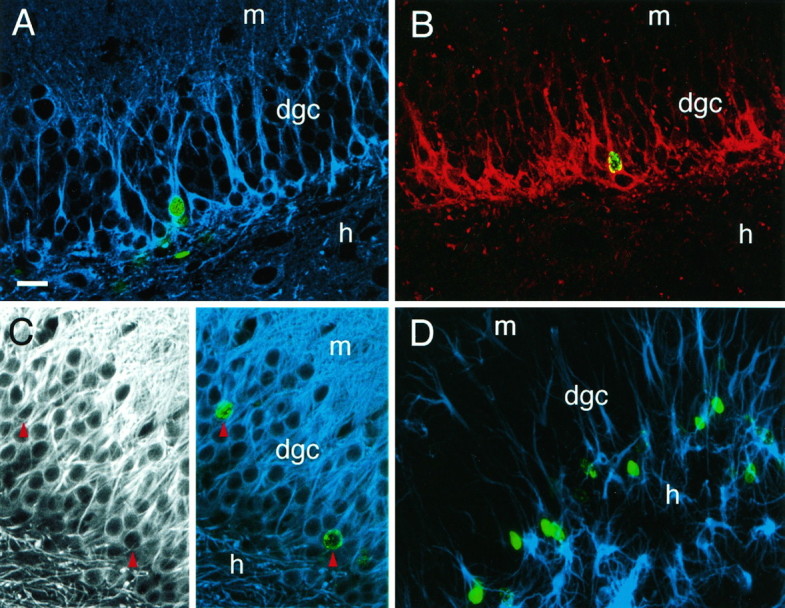
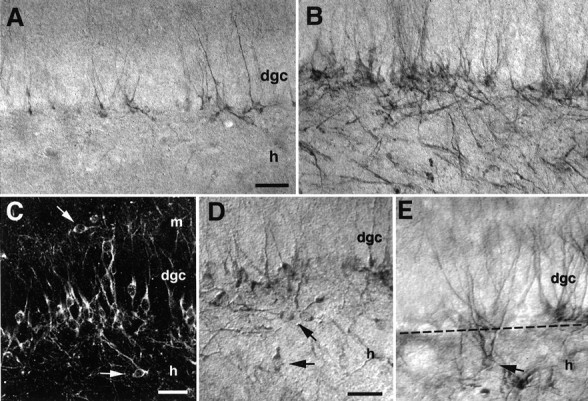
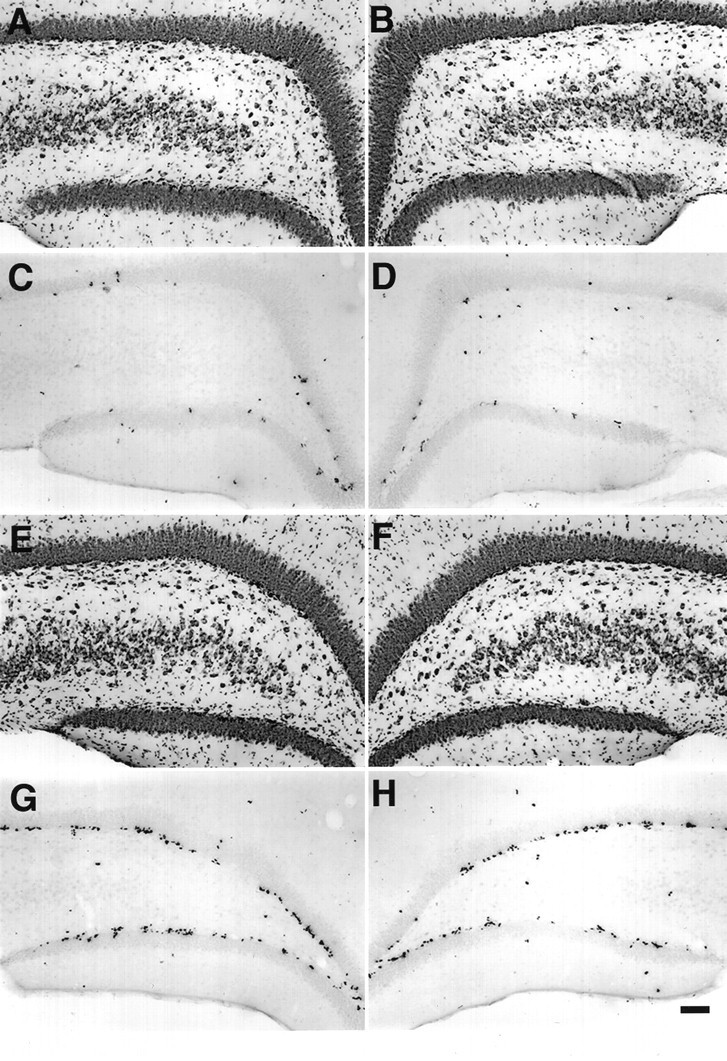
The dentate granule cell layer of the rodent hippocampal formation has the distinctive property of ongoing neurogenesis that continues throughout adult life. In both human temporal lobe epilepsy and rodent models of limbic epilepsy, this same neuronal population undergoes extensive remodeling, including reorganization of mossy fibers, dispersion of the granule cell layer, and the appearance of granule cells in ectopic locations within the dentate gyrus. The mechanistic basis of these abnormalities, as well as their potential relationship to dentate granule cell neurogenesis, is unknown. We used a systemic chemoconvulsant model of temporal lobe epilepsy and bromodeoxyuridine (BrdU) labeling to investigate the effects of prolonged seizures on dentate granule cell neurogenesis in adult rats, and to examine the contribution of newly differentiated dentate granule cells to the network changes seen in this model. Pilocarpine-induced status epilepticus caused a dramatic and prolonged increase in cell proliferation in the dentate subgranular proliferative zone (SGZ), an area known to contain neuronal precursor cells. Colocalization of BrdU-immunolabeled cells with the neuron-specific markers turned on after division, 64 kDa, class III beta-tubulin, or microtubule-associated protein-2 showed that the vast majority of these mitotically active cells differentiated into neurons in the granule cell layer. Newly generated dentate granule cells also appeared in ectopic locations in the hilus and inner molecular layer of the dentate gyrus. Furthermore, developing granule cells projected axons aberrantly to both the CA3 pyramidal cell region and the dentate inner molecular layer. Induction of hippocampal seizure activity by perforant path stimulation resulted in an increase in SGZ mitotic activity similar to that seen with pilocarpine administration. These observations indicate that prolonged seizure discharges stimulate dentate granule cell neurogenesis, and that hippocampal network plasticity associated with epileptogenesis may arise from aberrant connections formed by newly born dentate granule cells.
Figures

References
-
- Altar CA, Baudry M. Systemic injection of kainic acid: gliosis in olfactory and limbic brain regions quantified with [3H]PK 11195 binding autoradiography. Exp Neurol. 1990;109:333–341. - PubMed
-
- Altman J, Das GD. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J Comp Neurol. 1965;124:319–335. - PubMed
-
- Altman J, Das GD. Postnatal neurogenesis in the guinea pig. Nature. 1967;214:1098–1101. - PubMed
-
- Babb TL, Kupfer WR, Pretorious JK, Crandall PH, Levesque MF. Synaptic reorganization by mossy fibers in human epileptic fascia dentata. Neuroscience. 1991;42:351–363. - PubMed
-
- Bayer SA, Yackel JW. Neurons in the rat dentate gyrus granular layer substantially increase during juvenile and adult life. Science. 1982;216:890–892. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous