

Expression, stability, and membrane integration of truncation mutants of bovine rhodopsin
- PMID: 9144173
- PMCID: PMC24614
- DOI: 10.1073/pnas.94.10.4966
Expression, stability, and membrane integration of truncation mutants of bovine rhodopsin
Abstract
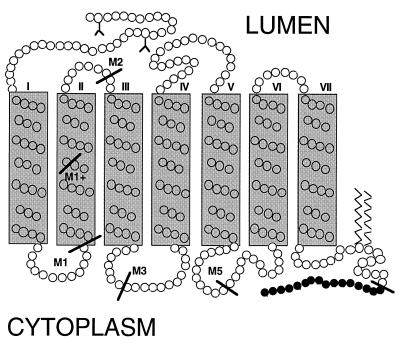
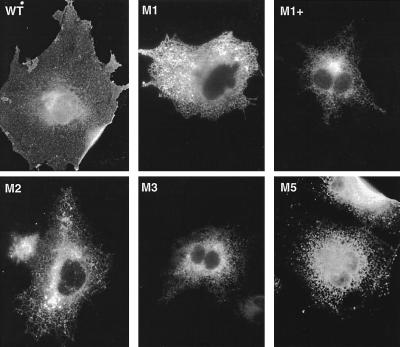
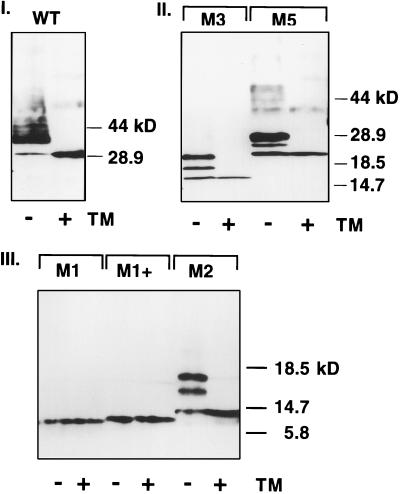
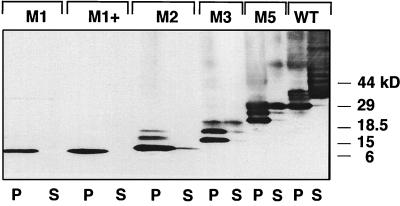
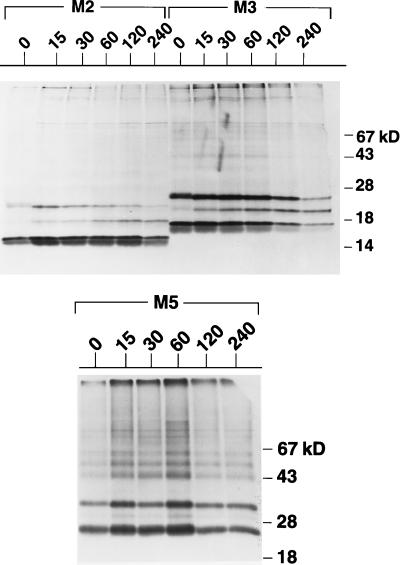
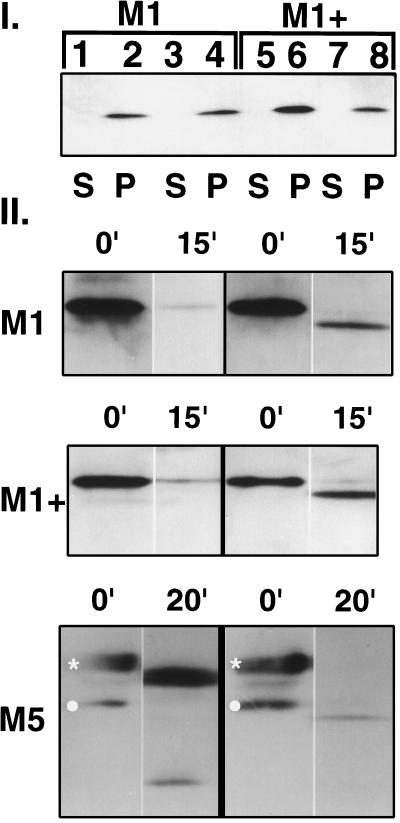
Premature termination of protein synthesis by nonsense mutations is at the molecular origin of a number of inherited disorders in the family of G protein-coupled seven-helix receptor proteins. To understand how such truncated polypeptides are processed by the cell, we have carried out COS-1 cell expression studies of mutants of bovine rhodopsin truncated at the first 1, 1.5, 2, 3, or 5 transmembrane segments (TMS) of the seven present in wild-type opsin. Our experiments show that successful completion of different stages in the cellular processing of the protein [membrane insertion, N-linked glycosylation, stability to proteolytic degradation, and transport from the endoplasmic reticulum (ER) membrane] requires progressively longer lengths of the polypeptide chain. Thus, none of the truncations affected the ability of the polypeptides to be integral membrane proteins. C-terminal truncations that generated polypeptides with fewer than two TMS resulted in misorientation and prevented glycosylation at the N terminus, whereas truncations that generated polypeptides with fewer than five TMS greatly destabilized the protein. However, all of the truncations prevented exit of the polypeptide from the ER. We conclude that during the biogenesis of rhodopsin, proper integration into the ER membrane occurs only after the synthesis of at least two TMS is completed. Synthesis of the next three TMS confers a gradual increase in stability, whereas the presence of more than five TMS is necessary for exit from the ER.
Figures
References
-
- Coughlin S R. Curr Opin Cell Biol. 1994;6:191–197. - PubMed
-
- Spiegel A M. Annu Rev Physiol. 1996;58:143–170. - PubMed
-
- Pan Y, Metzenberg A, Das S, Jing B, Gitschier J. Nat Genet. 1992;2:103–106. - PubMed
-
- Jacobson S G, Kemp C M, Cideciyan A V, Macke J P, Sung C H, Nathans J. Invest Ophthalmol Visual Sci. 1994;35:2521–2534. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
